Wednesday, December 25, 2013
Power From Urine
1. Urine is put into an electrolytic cell, which separates out the hydrogen.
2. The hydrogen enters a water filter for purification, which then gets pushed into a gas cylinder.
3. The gas cylinder pushes hydrogen into a cylinder of liquid borax, which is used to remove the moisture from the hydrogen.
4. This purified hydrogen gas is pushed into the generator.
Sunday, December 22, 2013
Saturday, December 21, 2013
Human Giants Of The Old Earth
This presentation was banned from ted talks because it was "debunked"
Jim Viera then easily debunked the debunk.
He provides data on some amazing discoveries including Ancient Giants of North America.
Well worth a watch.
This is a text of his explanation of suppression of this subject.
He also only claims that the "giants" were 7 to 11 feet. Probably more around 7 feet.
Stonemason and historical detective, Jim Vieira, discussed the ceremonial stone structures of North America as well as how the archaeological evidence for giants in America has been covered up.” There are structures of earth and stone, thousands of years old, in every part of the United States,” Vieira marveled. While he has found over 10,000 stone mounds around Massachusetts alone, he lamented that many great works were dismantled during colonial times as settlers used the materials for their roads and bridges. For instance, Vieira cited an archaeologist’s report of a 55-foot high stone pyramid in Illinois that had been plundered by colonists, who removed “tens of thousands of cartloads of stone.” Regarding giants in America, Vieira explained that, while searching through old town histories in Massachusetts for information on stone structures, he stumbled upon remarkable accounts of anomalous human skeletons being discovered. According to the reports from the time, the skeletons ranged from 7 to 11 feet tall and had skulls which sported double rows of teeth. “There are thousands of these accounts,” Vieira said, “by doctors, scientists, anthropologists, archaeologists, farmers and over decades and decades of time all around the country.” These skeletons, he added, were often found within the mysterious stone mounds and, sometimes, were sometimes buried amongst great treasures.
Vieira attributed the suppression of information on the ancient American giants to the Smithsonian Institute, specifically their first director of anthropology Ales Hrdlicka. Described by Vieira as a “pre-Nazi eugenicist,” Hrdlicka allegedly censored reports of giants and marginalized them from the historic record in order to depict Native Americans as “unsophisticated and uninspiring.” Additionally, Vieira reported that there are many accounts of Smithsonian representatives traveling around the country to where these anomalous remains were discovered and “taking the giant skeletons away, never to be seen again.” Ultimately, he contended that the cover up continues to this day and claimed that a video presentation on his findings was pulled from the Internet because it threatened the scientific orthodoxy.
Biography:
Stonemason and historical detective, Jim Vieira, studies the ancient ceremonial structures of North America including mounds, walls and star cities, as well as the archeological evidence for giants that existed in ancient America.

Thursday, December 19, 2013
Monday, December 9, 2013
The VASIMR Plasma Rocket: Bridging the Gap in Space Travel
Source

Plasma rocket technology was first introduced to the scientific community in 1977 by Franklin Chang Díaz, a Costa Rican scientist and astronaut. The idea has been developed since then and is now at the stage where it can be implemented into future projects. The technology allows for considerably faster space travel than what the technologies currently employed by organizations such as NASA can do.
The new plasma rocket is able to heat the argon gas, creating the plasma which is then focused out electromagnetically to give the rocket its propulsion. A shield protects the machinery from what is known as electrode erosion, which is a major cause of general wear and tear on a plasma rocket.
The thing that excites most scientists about the applications of the new rocket is a mission to Mars. The main problems in previous missions to Mars were that it would take too much fuel to allow the spaceship to reach the red planet and make a return journey. Deep space is another area that opens up to space explorers with the abilities of the plasma rocket. Travels to the areas of space that have remained inaccessible are back in the realm of possibility for scientists. Space debris removal as well as in-space resource recovery are also said to be possible with the new plasma rocket technology. Additionally, the magnetic field created by the rocket is thought to create a protective shell for the spaceship, which would help protect it against the radiation in space.
The 39-Day Mission to Mars
The rocket will allow for a mission to Mars with a travel period of just 39 days, which is almost 6 times faster than current methods. With speeds estimated at 35 miles a second, the rocket system will make quick work of the distance between Earth and Mars. NASA rates a new system on a scale of one to ten based on its readiness to be deployed. The VASIMR system is at a six currently, which means that it is ready to be tested in space. It would seem that it won’t be much longer before the new rocket system is employed in all space missions.
Plasma rocket technology was first introduced to the scientific community in 1977 by Franklin Chang Díaz, a Costa Rican scientist and astronaut. The idea has been developed since then and is now at the stage where it can be implemented into future projects. The technology allows for considerably faster space travel than what the technologies currently employed by organizations such as NASA can do.
What is the VASIMR Plasma Rocket?
VASIMR stands for Variable Specific Impulse Magnetoplasma Rocket, which makes use of argon gas (one of the most stable gasses known to man) and a renewable source of energy found in space, radio waves in the form of light. The main difference with this type of rocket is being able to use mostly renewable energy in the propulsion system, which gives the rocket a greater lifespan than similar, modern-day rocket technologies. The plasma technology has multiple applications such as the cleaning and coating of surfaces in a plasma coating system at nano-level. The uses of plasma, the 4th state of substances, are just being touched on now with recent advances in science.
VASIMR stands for Variable Specific Impulse Magnetoplasma Rocket, which makes use of argon gas (one of the most stable gasses known to man) and a renewable source of energy found in space, radio waves in the form of light. The main difference with this type of rocket is being able to use mostly renewable energy in the propulsion system, which gives the rocket a greater lifespan than similar, modern-day rocket technologies. The plasma technology has multiple applications such as the cleaning and coating of surfaces in a plasma coating system at nano-level. The uses of plasma, the 4th state of substances, are just being touched on now with recent advances in science.
VASIMR’s Impact on Space Travel
Over the last 36 years, the various shortcomings of the propulsion system have been ironed out. Problems such as wear and tear as well as the total velocity it can achieve have been improved dramatically. This new form of thrust is said to cut space mission times by up to six times. With the use of the plasma rocket, it will be possible to make quick surveys to other planets or even asteroids that may be on a collision course with Earth.
Over the last 36 years, the various shortcomings of the propulsion system have been ironed out. Problems such as wear and tear as well as the total velocity it can achieve have been improved dramatically. This new form of thrust is said to cut space mission times by up to six times. With the use of the plasma rocket, it will be possible to make quick surveys to other planets or even asteroids that may be on a collision course with Earth.
The 39-Day Mission to Mars
The rocket will allow for a mission to Mars with a travel period of just 39 days, which is almost 6 times faster than current methods. With speeds estimated at 35 miles a second, the rocket system will make quick work of the distance between Earth and Mars. NASA rates a new system on a scale of one to ten based on its readiness to be deployed. The VASIMR system is at a six currently, which means that it is ready to be tested in space. It would seem that it won’t be much longer before the new rocket system is employed in all space missions.
Wednesday, November 13, 2013
UFOs - The Black Knight Satellite
Published on Jun 1, 2013
What is the "Black Knight" satellite? It is a mysterious satellite, of unknown origin, discovered in 1960 which shadowed Sputnik. It is believed to have been of extraterrestrial origin, and signaled back old radio waves from the 1920s and 1930s before it disappeared. In short wave patterns analyzed by astronomer Duncan Lunan, it revealed its origin as Epsilon Boötes (or the star system as it was 13,000 years ago).
Based on reports, The Black Knight satellite has returned. Photos of the Black Knight satellite, and its sudden reappearance around the orbit of Earth, were available on NASA's website up until January 2013, until they disappeared. This video contains the photos from NASA's archives.
Based on reports, The Black Knight satellite has returned. Photos of the Black Knight satellite, and its sudden reappearance around the orbit of Earth, were available on NASA's website up until January 2013, until they disappeared. This video contains the photos from NASA's archives.



In March of 1960, another mysterious satellite was discovered orbiting earth. This mystery satellite was in polar orbit feat that neither the US nor Russian were capable of at that time. The soviet satellites were inclined at 65 degree angle with the equator. It was calculated that the weight object to about 15 tons, which is far too heavy for rocket of that era. It also traveled twice as fast as any known satellite at the time. The object was sighted and studied by many of the astronomers across the country. This is what Robert L. Johnson, director of the Adler Planetarium had to say about the object,
“ ‘THE OBJECT,’ JUDSON SAID, ‘DOESN’T EVEN HAVE THE DECENCY TO MAINTAIN A REGULAR SCHEDULE,LIKE ANY OTHER HEAVENLY OR MAN MADE OBJECT HE’S EVER SEEN.’ ‘WE DON’T KNOW WHEN TO WATCH FOR IT.’ he says.” It appears some nights, and some nights it doesn’t.
It was then photographed by the Grumman Aircraft Corporation.
The military attached so much importance to this mystery satellite that they created special committee to gather information and study the mystery satellite. The committee’s findings were kept secret and the sightings were forgotten. That is still HAM operator intercepted a strange signal from this mystery satellite. The signal was later studied by Scottish astronomer Duncan Lunan. The decoded massages show a star map centered on Epsilon Boštes as seen from the earth. . . years ago.

In 1953, four years before the U.S.S.R. launched Sputnik I, a the blip of unknown origin was sighted by Dr Lincoln La Paz of the University of New Mexico. As more reports of sightings trickled in from around the world, the U.S. Department of Defense appointed distinguished astronomer Clyde W. Tombaugh to run a search for the mystery object. He had discovered Pluto in 1930. The blip became known as black knight.
The Pentagon never formally released the results of Dr Tombaugh’s study. And no more was heard about the object until December, 1957, when Dr Luis Corralos of the Communications Ministry in Venezuela photographed it. The first modern satellite, Sputnik I, had been launched two months earlier. Dr Corralos was taking pictures of Sputnik II as it passed over Caracas.
In 1957 black knight the blip of unknown origin was discovered in 1960 by a satellite. It was detected shadowing the sputnik 1 craft It was in a polar orbit, something that neither the Americans or Soviets were capable of at the time. This object was dubbed “The Black Knight. It was several times larger and several times heavier than anything capable of being launched with 1960 rockets. It shouldn’t have been there, but it was.
Time Magazine published this article on Monday, March 7th, 1960;
http://www.time.com/time/magazine/article/0,9171,894745,00.html
1960, February – North American Air Defense System (NORAD predecessor) detects object with a mass of 15 metric tons in Polar orbit. It is several times larger than anything the US or Soviets are capable of launching into orbit. The object, dubbed “Black Knight,” is tracked for three weeks and abruptly disappears. Ham radio operators once again report LDE phenomenon during this period.

Read more
Friday, October 18, 2013
The World's Tallest Tree Is Hiding Somewhere In California
Source
by ROBERT KRULWICH
April 08, 201110:34 AM
Woah, this is one very tall tree. Nine years ago, it was the tallest known plant in the world. (See if you can spot the three people, blue shirted, hanging at different points on the trunk).

Credit: Photo by James Balog
It's 369 feet high. That's about twice the size of the Statue of Liberty (minus the foundation). I like this tree. The people who discovered it have never revealed its true location, which is somewhere in Humboldt Redwoods State Park. And though it's got a nickname, "Stratosphere Giant," it is no longer the giant. It's been trumped.
After its short four-year reign as World's Tallest, two hikers, Chris Atkins and Michael Taylor, were deep in another section of another park, Redwood National Park (purchased in 1978 during the Carter administration) when they came across a new stand of trees, taller than anyone had ever seen before. The tallest of the tall is 379 feet 4 inches, 10 feet taller than the Giant. It's now called "Hyperion."
We have the precise measurements because after Chris and Michael announced their discovery, a team of scientists, led by Humboldt State University ecologist Steve Sillett, climbed to the top of the tree and dropped a tape down to the ground. Some things are still that simple. Steve's colleague, Jim Spickler (check out his biceps! scary), repeated the climb and brought a camera, so we can go with him. This video, which comes with dramatic music in all the right places, is, to use a much overused word, but I'll use it anyway..."awesome":
Once again, no one is telling where the new champ is located. People will want to see it, photograph it, climb it, carve little souvenirs out of it. Trees, says Steve Sillett, are not like people. They "cannot run away from paparazzi." He asked us not to even mention the nickname, "Hyperion," but I figure the nickname gives away nothing. Anyway, the Who's Biggest title keeps changing.
For example, if you like wood, Hyperion is not the biggest tree in the world. By volume, there are bigger hunks of living wood in the California forest. A REALLY big one, by volume, is this guy ...

Del Norte TitanDavid Mickelson/groveoftitans.com
According to Nalini Nadkarni, a professor at Evergreen State College, there is so much wood in this tree, called Del Norte Titan (discovered by the same Steve Sillett who climbed Hyperion), "its mass is equivalent to 15 adult blue whales, the largest animal on earth. Each year, this tree produces enough new wood to make a 90-foot-tall tree with a trunk 12 inches in diameter. If all of Del Norte Titan were cut into boards one foot wide, 12 feet long and one inch thick, the line of planks laid end to end would stretch over a hundred miles and could build 120 average-sized houses."
But nobody's going to be chopping up Del Norte Titan. Donald Trump can drool all he likes, but these 15 blue whales' worth of lumber will stay vertical somewhere in Jedediah Smith Redwoods State Park in California, breathing like trees do, pulling CO2 out of the air and giving us back oxygen.
That's the real gift of trees. Yes, we live under roofs made of wood, surround ourselves with woody things, but — basically, what we breathe out, they breathe in.
As the poet Bill Yake writes:
The photo collage of the Stratosphere Giant was created by photographer James Balog; Nalini Nadkarni's book on trees is "Between Earth and Sky: Our Intimate Connections to Trees (University of California Press, 2008); Bill Yake's poem is called "Inside Out."
by ROBERT KRULWICH
April 08, 201110:34 AM
Woah, this is one very tall tree. Nine years ago, it was the tallest known plant in the world. (See if you can spot the three people, blue shirted, hanging at different points on the trunk).
Credit: Photo by James Balog
It's 369 feet high. That's about twice the size of the Statue of Liberty (minus the foundation). I like this tree. The people who discovered it have never revealed its true location, which is somewhere in Humboldt Redwoods State Park. And though it's got a nickname, "Stratosphere Giant," it is no longer the giant. It's been trumped.
After its short four-year reign as World's Tallest, two hikers, Chris Atkins and Michael Taylor, were deep in another section of another park, Redwood National Park (purchased in 1978 during the Carter administration) when they came across a new stand of trees, taller than anyone had ever seen before. The tallest of the tall is 379 feet 4 inches, 10 feet taller than the Giant. It's now called "Hyperion."
We have the precise measurements because after Chris and Michael announced their discovery, a team of scientists, led by Humboldt State University ecologist Steve Sillett, climbed to the top of the tree and dropped a tape down to the ground. Some things are still that simple. Steve's colleague, Jim Spickler (check out his biceps! scary), repeated the climb and brought a camera, so we can go with him. This video, which comes with dramatic music in all the right places, is, to use a much overused word, but I'll use it anyway..."awesome":
Once again, no one is telling where the new champ is located. People will want to see it, photograph it, climb it, carve little souvenirs out of it. Trees, says Steve Sillett, are not like people. They "cannot run away from paparazzi." He asked us not to even mention the nickname, "Hyperion," but I figure the nickname gives away nothing. Anyway, the Who's Biggest title keeps changing.
For example, if you like wood, Hyperion is not the biggest tree in the world. By volume, there are bigger hunks of living wood in the California forest. A REALLY big one, by volume, is this guy ...
Del Norte TitanDavid Mickelson/groveoftitans.com
According to Nalini Nadkarni, a professor at Evergreen State College, there is so much wood in this tree, called Del Norte Titan (discovered by the same Steve Sillett who climbed Hyperion), "its mass is equivalent to 15 adult blue whales, the largest animal on earth. Each year, this tree produces enough new wood to make a 90-foot-tall tree with a trunk 12 inches in diameter. If all of Del Norte Titan were cut into boards one foot wide, 12 feet long and one inch thick, the line of planks laid end to end would stretch over a hundred miles and could build 120 average-sized houses."
But nobody's going to be chopping up Del Norte Titan. Donald Trump can drool all he likes, but these 15 blue whales' worth of lumber will stay vertical somewhere in Jedediah Smith Redwoods State Park in California, breathing like trees do, pulling CO2 out of the air and giving us back oxygen.
That's the real gift of trees. Yes, we live under roofs made of wood, surround ourselves with woody things, but — basically, what we breathe out, they breathe in.
As the poet Bill Yake writes:
Trees are our lungs turned inside out
& inhale our visible chilled breath.
Our lungs are trees turned inside out
& inhale their clear exhalations.
Some things are still that simple.
The photo collage of the Stratosphere Giant was created by photographer James Balog; Nalini Nadkarni's book on trees is "Between Earth and Sky: Our Intimate Connections to Trees (University of California Press, 2008); Bill Yake's poem is called "Inside Out."
Thursday, October 3, 2013
Friday, September 13, 2013
Tuesday, September 10, 2013
5 Crazy New Man-Made Materials That Will Shape The Future

Forget Mother Nature: when it comes to all matters matter, the sheer ingenuity of the human mind can give rise to some of the most insane — and useful — new materials you’ve ever encountered. Here are five crazy new man-made materials whose uses could be practically limitless.
Aluminium Bubble Wrap

Imagine your favourite packing-based stress reliever — made of metal. OK, metallic bubble wrap might not be quite as easy to pop, but it could be a hell of a lot more useful. A team of engineers from North Carolina State University have developed a new form of aluminium bubble wrap, which they claim could revolutionise packaging and protective equipment.
The scientists take a thin sheet of aluminium, then use a studded roller to form small indents in the sheet. Unlike its polyethylene counterpart, these voids are then filled with a foamed material like calcium carbonate, before being sealed with another flat sheet of metal. The result: a series of bubbles that absorb masses of energy, weigh 30 per cent less than regular sheet metal, and yet are nearly 50 times stronger. It’s easy to make, not too expensive — and could soon be used in everything from shipping containers for fragile goods to bike helmets. Just don’t be tempted to try and pop it.
Titanium Foam

Forget expanded polystyrene and spongey elastomers: the foam you want to get your hands on is made out of titanium. By saturating a humble polyurethane foam with a solution of titanium powder and binding agents, it’s possible to force the metal to cling to the shape of the simple foam and then vaporise the underlying structure away. The result is a titanium lattice in the shape of the original foam, which can be heat-treated to tweak its material properties.
The exact properties depend on the porosity of the foam, but the results are strong and — most importantly — incredibly light. In fact, the material is just perfect for replacing bone: it has incredibly similar mechanical properties and, because it’s porous, new bone can grow and around its structure, truly integrating the implant within the skeleton. Anything that gets us that much closer to a real-life Wolverine is ok in our book.
Graphene Aerogel

If the phrase graphene aerogel sounds to you like someone combined the two buzziest of materials buzzwords then… you wouldn’t be far wrong. In fact, this graphene aerogel snatched the title of the world’s lightest material just a few of months ago — with a density lower than that of helium and just twice that of hydrogen at 0.16 mg/cm3. This stuff practically floats.
The material was actually created using a new technique which involves freeze-drying solutions of carbon nanotubes and graphene to create a kind of carbon sponge. The resulting material is both strong and elastic, as well as incredibly light; it can even absorb up to 900 times its own weight in oil. When — or if — it becomes affordable, that means it could be used to mop up massive oil spills with ease, or even as an incredibly efficient version of humble old insulation.
Artificial Spider Silk

Silk is Nature’s very own homespun wonder material but it’s tough to make in bulk — which is why a Japanese startup called Spiber has been working out how to produce it synthetically. It’s managed to decode the gene responsible for the production of fibroin in spiders, which is they key protein used to create the super-strong strands of silk.
Having cracked that key component, the company has gone on to create bioengineered bacteria that can make silk extremely quickly — and the company can create a new type of silk in just 10 days, from scratch to to finished product. The bacteria feed on sugar, salt and other micronutrients, and quickly produce the silk protein — which is turned into a fine powder, spun and processed to create fibres, composites, solid block… anything. A single gram of fibroin produces 5.6 miles of silk, and by 2015 the company hopes to be produce 10 metric tons of the stuff. That’s a lot of silk, without a spider in sight.
Molecular Superglue

If you’ve ever stuck your fingers together with superglue, you know pain — but imagine sticking them together with glue that bonds materials at the molecular level: that’s real pain. In fact, a team of researchers from the University of Oxford has created a molecular glue inspired by Streptococcus pyogenes — the flesh-eating bacteria.
They considered a single protein from the bacterium — the one it uses to bind to human cells — and from there developed a molecular glue which forms covenant bonds when it comes into contact with a partner protein. The bonds it forms are so strong that, when researchers tested a sample, the equipment used to measure the strength broke before the glue did. All that remains is to develop ways of incorporating the proteins into other molecular structures in order to create insanely strong, selective glues.
Friday, August 9, 2013
Strange Sea Creature Carcass
An Iranian Navy Ship came across this creature in the Persian Gulf when they noticed a terrible smell








Saturday, July 6, 2013
Friday, July 5, 2013
The Hybrid Hypothesis
Source
BY EUGENE M. MCCARTHY, PHD GENETICS
This article is a bit different from others that have appeared on this site, in that it's about some of the findings of my own research. I'm a geneticist whose work focuses on hybrids and, particularly, the role of hybridization in the evolutionary process. Here, I report certain facts, which seem to me to indicate that human origins can be traced to hybridization, specifically to hybridization involving the chimpanzee (but not the kind of hybridization you might suppose!). You can access detailed and documented discussions supporting this claim from links on this page. But the basic reasoning is summarized here, without a lot of citations and footnotes.
And why might one suppose that humans are backcross hybrids of the sort just described? Well, the most obvious reason is that humans are highly similar to chimpanzees at the genetic level, closer than they are to any other animal. If we were descended from F1 hybrids without any backcrossing we would be about halfway, genetically speaking, between chimpanzees and whatever organism was the other parent. But we're not. Genetically, we're close to chimpanzees, and yet we have many physical traits that distinguish us from chimpanzees. This exactly fits the backcross hypothesis.
Primatologist Adolph Schultz (1973), however, flatly contradicts Darwin, saying that "clearly pointed ears, commonly called `satyr ears,' are among monkeys typical for only macaques and baboons and do not occur in any hominoids [great apes], not even in the early stages of development. There is no justification, therefore, to interpret the occasional `Darwinian tubercles' on human ears as an atavistic manifestation of ancestral pointed ears."20 But Schultz has not, perhaps, taken into consideration the pointed ears of swine.
As Cena et al. (101.9,521) observe, "Hair densities [of animal coats] range from the sparse residual covering on man and the pig with 10-100 hairs per cm², to [the] dense coats of species such as the fox and rabbit with about 4,000 per cm²." In wildSus scrofa, according to Haltenorth, the density of hair coverage varies from "sparse to thick," depending on the specimen or variety in question.23 For example, the hair of the modern day wild variety of Sus scrofa present in Sudan (S. s. senaarensis) is quite sparse.24
More significantly, it brings the base of the flexible portion of the spinal column into a position directly above the hip joints (when viewed from the side). The force applied to the pelvis by the weight of the upper body is directed straight downward through the hip joints and does not tend to rotate the pelvis around those joints. When an ape is fully erect, a vertical line passing through the base of the spine falls behind the hip joints so that a rearward twisting torque is applied to the pelvis. This torque must be countered by constant muscular exertion.
Moreover, the figures above clearly show that human cervicals are generally more similar to a pig's than to those of an ape.
Under the assumption that humans stand on a "higher" rung of the evolutionary ladder, chimpanzees should have a longer and more pliable "tail" than do humans. But, in fact, the human coccyx is not fused, but movable — especially in females, where it bends backward when they are giving birth.27
Perhaps we should look elsewhere for an explanation. The sacrum of a pig is curved on its inner side much like that of a human being (see illustration above). Obviously, pigs have tails, albeit short ones. If Homo is a hybrid of ape and pig, we expect the human sacrum to be curved and the coccyx to be longer and more flexible than an ape's. The human pelvis is peculiar in many respects. According to Adolph Schultz,
"In hominids, both condyles show a distinct elliptical shape, indicating a specialization for maximum cartilage contact in the knee joint only during full extension of the lower limb. In [primate] quadrupeds, on the other hand, the condyles show no such specialization to one position, being essentially circular in cross-sectional outline" (Lovejoy46). Nevertheless, many non-primate quadrupeds do, in fact, have elliptical femoral condyles. Among them are most of the domestic animals: cows, sheep, horses, dogs — and pigs (see illustration below).47 We have no reason, then, to think that human elliptical condyles represent an adaptation aiding in bipedal locomotion.
In Homo this downwardly directed, spike-like process is reduced to little more than a nub. In pigs it is so short as to be nearly nonexistent.55
Shrewsbury and Johnson state that "the distinguishing features of the human distal phalanx are the broad spade-like tuberosity with proximal projecting spine and the wide diaphysis, which is concave palmarly to create an ungual fossa. These features are not seen in primates such as the monkey and gorilla."64 This distinction, which was also present the various extinct hominids (395.5,539,541), has been explained as an adaptation facilitating the manipulation of objects with the fingertips. If such is the case, why should these processes also be found on the tips of our toes? Do these hoof-like ungual tuberosities actually reflect a relationship between humans and ungulates like the pig? That is, are they vestiges of ancestral hooves?
BY EUGENE M. MCCARTHY, PHD GENETICS
 | |
| Chimpanzee (Pan troglodytes) | |
| "The scientist has a lot of experience with ignorance and doubt and uncertainty, and this experience is of very great importance, I think. When a scientist doesn’t know the answer to a problem, he is ignorant. When he has a hunch as to what the result is, he is uncertain. And when he is pretty darn sure of what the result is going to be, he is in some doubt. We have found it of paramount importance that in order to progress we must recognize the ignorance and leave room for doubt. Scientific knowledge is a body of statements of varying degrees of certainty — some most unsure, some nearly sure, none absolutely certain."
— Richard Feynman
|
Rationale
So why do I think humans are hybrids? Well, first of all, I've had a different experience from most other people. I've spent most of my life (the last thirty years) studying hybrids, particularly avian and mammalian hybrids. I've read thousands, really tens of thousands, of reports describing them. And this experience has dispelled some mistaken ideas I once had about hybrids, notions that I notice many other people continue to take for granted.
For example, one widespread, but erroneous belief that keeps a lot of people from even considering the possibility that humans might be of hybrid origin is the notion that all hybrids are sterile. This assertion, though I've heard lots of people say it, is absolutely false. For instance, in reviewing the reports I collected for my book on hybridization in birds (Handbook of Avian Hybrids of the World,Oxford University Press, 2006), which documents some 5,000 different kinds of hybrid crosses among birds, I found that those producing partially fertile hybrids are about eight times as common as crosses known to produce sterile ones. So the usual result is areduction in fertility, not absolute sterility. My current work documenting hybridization among mammals shows that partially fertile natural hybrids are common, too, in Class Mammalia. And yet, it seems most people base their ideas of hybrids on the common mule (horse x ass), which is an exceptionally sterile hybrid, and not at all representative of hybrids as a whole.
A second "fact" that might make it seem impossible for humans to have had a hybrid origin is the equally erroneous notion that hybrids, especially successful hybrids, do not occur in a state of nature. A third is the mistaken idea that only plants hybridize and never animals. In fact, however, natural, viable, fertile animal hybrids are abundant. A wide variety of such hybrids occur on an ongoing basis (read a detailed discussion documenting these facts). For example, of the 5,000 different types of hybrid crosses listed in my book on hybridization in birds, approximately half are known to occur in a natural setting (download a PowerPoint presentation summarizing data on hybridization in birds). My current research indicates a comparable rate for mammals.
Sequence data And I must now emphasize a fact that I, as a geneticist, find somewhat disappointing: Nucleotide sequence data can't be used to identify later-generation backcross hybrids derived from several repeated generations of backcrossing (for a full explanation of this fact, see the green sidebar at far right). Instead, as is the case with other later-generation backcross hybrids, the most revealing data is of an anatomical and/or physiological nature. (For those readers who do not know, backcross hybrids are produced when hybrids from a first cross mate with either of the two types of parents that produced them. When the resulting progeny mate again with the same parental type, the result is the second backcross generation, and so forth.)
Human infertility. Another observation that appears significant in connection with the hypothesis under consideration is that it has been well known for decades that human sperm is abnormal in comparison with that of the typical mammal. Human spermatozoa are not of one uniform type as in the vast majority of all other types of animals. Moreover, human sperm is not merely abnormal in appearance — a high percentage of human spermatozoa are actually dysfunctional. These and other facts demonstrate that human fertility is low in comparison with that of other mammals (for detailed documentation of this fact see the article Evidence of Human Infertility). Infertility and sperm abnormalities are characteristic of hybrids. So this finding suggests that it's reasonable to suppose, at least for the sake of argument, that humans might be of hybrid origin. It is also consistent with the idea that the hybridization in question was between two rather distinct and genetically incompatible types of animals, that is, it was a distant cross.
| A personal endorsement: "As a clinician and scientist with medical training it is a joy to find a theory so carefully and elegantly presented. My interest in the hybrid nature of modern man led me to Eugene McCarthy's website and lifework. What a revelation! Surprising and shocking. Such is the nature of truth sometimes. Life will never be seen in the same way after reading this work." Dr Chris Millar Ballarat, Victoria, Australia |
Methodology The chimpanzee is plausible in the role of one of parents that crossed to produce the human race because they are generally recognized as being closest to humans in terms of their genetics (here, I use the term (chimpanzeeloosely to refer to either the common chimpanzee or to the bonobo, also known as the pygmy chimpanzee; the specific roles of these two rather similar apes within the context of the present hypothesis will be explained in a subsequent section). But then the question arises: If an ancient cross between the chimpanzee and some parental form "X" produced the first humans, then what was that parent? Does it still exist? What was it like?
As the reader might imagine, if the assumption is correct that one of our parents is the chimpanzee, then it should be possible actually to identify the other parent as well. A hybrid combines traits otherwise seen only separately in the two parental forms from which it is derived, and it is typically intermediate to those parents with respect to a wide range of characters. Naturalists routinely use these facts to identify the parents of hybrids of unknown origin, even backcross hybrids.
First they posit a particular type of organism as similar to the putative hybrid (in the present case, this organism is the chimpanzee). They then list traits distinguishing the hybrid from the hypothesized parent, and this list of distinguishing traits will describe the second parent. A detailed analysis of such a triad will often establish the parentage of the hybrid. The traits in question in such studies are generally anatomical, not genetic. DNA evidence is used in only a very small percentage such identifications (and even then, not to identify backcross hybrids), and yet firm conclusions can generally be reached.
So in the specific case of humans, if the two assumptions made thus far are correct (i.e., (1) that humans actually are hybrids, and (2) that the chimpanzee actually is one of our two parents), then a list of traits distinguishing human beings from chimpanzees should describe the other parent involved in the cross. And by applying this sort of methodology, I have in fact succeeded in narrowing things down to a particular candidate. Keep reading and I'll explain.
| Teiresias: To you, I am mad; but not to your parents. Oedipus: Wait! My parents? Who are my parents? |
| —Sophocles, Oedipus Tyrannus |
 |
Moreover, in mammalian hybrid crosses, the male hybrids are usually more sterile than are the females. In a commercial context, this fact means that livestock breeders typically backcross F1 hybrids of the fertile sex back to one parent or the other. They do not, as a rule, produce new breeds by breeding the first cross hybrids among themselves. Often, even after a backcross, only the females are fertile among the resulting hybrids. So repeated backcrossing is typical. Commonly there are two or more generations of backcrossing before fertile hybrids of both sexes are obtained and the new breed can be maintained via matings among the hybrids themselves. More backcrossing tends to be necessary in cases where the parents participating in the original cross are more distantly related.
Traits distinguishing humans from other primates
| A reader's comment: "Your conjecture is not unlike trying to reverse engineer a human being. Logically it all makes a good argument, down to the detailed level you've taken it to. I imagine that working with hybrids you HAVE to do that - even in cases where you may not think so. Logically your arguments make a lot of sense. And the corollaries and ramifications all seem to come true. I am impressed, frankly." |
Many characteristics that clearly distinguish humans from chimps have been noted by various authorities over the years. The task of preliminarily identifying a likely pair of parents, then, is straightforward: Make a list of all such characteristics and then see if it describes a particular animal. One fact, however, suggests the need for an open mind: as it turns out, many features that distinguish humans from chimpanzees also distinguish them from all other primates. Features found in human beings, but not in other primates, cannot be accounted for by hybridization of a primate with some other primate. If hybridization is to explain such features, the cross will have to be between a chimpanzee and a nonprimate — an unusual, distant cross to create an unusual creature.
The fact that even modern-day humans are relatively infertile may be significant in this connection. If a hybrid population does not die out altogether, it will tend to improve in fertility with each passing generation under the pressure of natural selection. Fossils indicate that we have had at least 200,000 years to recover our fertility since the time that the first modern humans (Homo sapiens) appeared. The earliest creatures generally recognized as human ancestors (Ardipithecus, Orrorin) date to about six million years ago. So our fertility has had a very long time to recover. If we have been improving for thousands of generations and still show obvious symptoms of sterility (see previous section), then our earliest human ancestors, if they were hybrids, must have suffered from an infertility that was quite severe. This line of reasoning, too, suggests that the chimpanzee might have produced Homo sapiensby crossing with a genetically incompatible mate, possibly even one outside the primate order.
For the present, I ask the reader to reserve judgment concerning the plausibility of such a cross. I'm an expert on hybrids and I can assure you that our understanding of hybridization at the molecular level is still far too vague to rule out the idea of a chimpanzee crossing with a nonprimate. Anyone who speaks with certainty on this point speaks from prejudice, not knowledge. No systematic attempts to cross distantly related mammals have been reported. However, in the only animal class (Pisces) where distant crosses have been investigated scientifically, the results have been surprisingly successful (399.6, 399.7, 399.8). In fact, there seems to be absolutely nothing to support the idea that interordinal crosses (such as a cross between a primate and a nonprimate) are impossible, except what Thomas Huxley termed "the general and natural belief that deliberate and reiterated assertions must have some foundation."
Besides, to deny that interordinal mammalian crosses are possible would be to draw, at the outset of our investigation, a definite conclusion concerning the very hypothesis that we have chosen to investigate. Obviously, if humans were the product of such a cross, then such crosses would, in fact, be possible. We cannot tell, simply by supposing, whether such a thing is possible — we have to look at data.
The Other Parent
Let's begin, then, by considering the list in the sidebar at right, which is a condensed list of traits distinguishing humans from chimpanzees — and all other nonhuman primates. Take the time to read this list and to consider what creature — of any kind — it might describe. Most of the items listed are of such an obscure nature that the reader might be hard pressed to say what animal might have them (only a specialist would be familiar with many of the terms listed, but all the necessary jargon will be defined and explained). For example, consider multipyramidal kidneys. It's a fact that humans have this trait, and that chimpanzees and other primates do not, but the average person on the street would probably have no idea what animals do have this feature.
Looking at a subset of the listed traits, however, it's clear that the other parent in this hypothetical cross that produced the first human would be an intelligent animal with a protrusive, cartilaginous nose, a thick layer of subcutaneous fat, short digits, and a naked skin. It would be terrestrial, not arboreal, and adaptable to a wide range of foods and environments. These traits may bring a particular creature to mind. In fact, a particular nonprimate does have, not only each of the few traits just mentioned, but every one of the many traits listed in th sidebar. Ask yourself: Is it likely that an animal unrelated to humans would possess so many of the "human" characteristics that distinguish us from primates? That is, could it be a mere coincidence? It's only my opinion, but I don't think so.
Of course, it must be admitted that two human traits do, at first, seem to pose a contradiction. The animal in question lacks a large brain and it is not bipedal. An analysis of the relevant anatomy, however, reveals that these two human features can be understood as synergistic (or heterotic) effects, resulting from the combination (in humans) of certain traits previously found only separately, in the two posited parent forms. (The origins of human bipedality is explained in terms of the the hybrid hypothesis in a subsequent section.Another section offers an explanation of the factors underlying human brain expansion and, therefore, accounts not only for the large size of the human brain itself, but also for certain distinctive features of the human skull that are, themselves, obvious consequences of brain expansion).
Nevertheless, even initially, these two flies in the theoretical ointment fail to obscure the remarkable fact that a single nonprimate has all of the simple, non-synergistic traits distinguishing humans from their primate kin. Such a finding is strongly consistent with the hypothesis that this particular animal once hybridized with the chimpanzee to produce the first humans. In a very simple manner, this assumption immediately accounts for a large number of facts that otherwise appear to be entirely unrelated.
| From a recent Twitter conversation with a biologist who says he's convinced by the argument presented in the Hybrid Hypothesis: | |
 |
What is this other animal that has all these traits? The answer is Sus scrofa, the ordinary pig. What are we to think of this fact? If we conclude that pigs did in fact cross with apes to produce the human race, then an avalanche of old ideas must crash to the earth. But, of course, the usual response to any new perspective is "That can't be right, because I don't already believe it." This is the very response that many people had when Darwin first proposed that humans might be descended from apes, an idea that was perceived as ridiculous, or even as subversive and dangerous. And yet, today this exact viewpoint is widely entertained. Its wide acceptance can be attributed primarily to the established fact that humans hold many traits in common with primates. That's what made it convincing. But perhaps Darwin told only half the story. We believe that humans are related to chimpanzees because humans share so many traits with chimpanzees. Is it not rational then also, if pigs have all the traits that distinguish humans from other primates, to suppose that humans are also related to pigs? Let us take it as our hypothesis, then, that humans are the product of ancient hybridization between pig and chimpanzee. Given the facts presented in the discussion of stabilization theory on this website, it seems highly likely that humans are hybrids of some kind.
This particular hypothesis concerning the nature of our parentage is, as we shall see, a fruitful one. For the present there's no need to make a definite decision on the matter, but certain lines of reasoning do suggest the idea should be taken seriously:
| A reader's comment: "Wow! I learned of this site and your pig-chimpanzee-hybrid paper only a few hours ago, and have been stuck here ever since. Fantastic work." |
- First of all, the notion is set forward strictly as a hypothesis. No claim whatever is made that it is actually a fact that humans somehow arose through hybridization of pigs with chimpanzees. In contrast, proponents of the idea that humans are closely related to apes (and not to pigs) often speak as if their case has been proved beyond doubt. But, of course, it has not. The wide acceptance of this idea may actually be due to the lack of any competitive theory. I merely propose an evaluation of two distinct hypotheses by the usual scientific criterion: The hypothesis less consistent with available data should be rejected.
- Even if we could identify some objective unit of measure for "distance" or "similarity" (which is not at all a straightforward problem), we would still expect some crosses to be more distant than others — that is, the various types of possible crosses would constitute a continuum. Many would be "close" and some would be "distant." But we would expect at least a rare few to be very distant. While these few might be rare, they might be among the most interesting, because they would offer an opportunity to obtain something radically different. Perhaps, it is only a subjective bias, but I believe that a human being, when taken as a whole, is radically different from a chimpanzee.
- On the other hand, if we first compare humans with nonmammals or invertebrates (e.g, crocodile, bullfrog, octopus, dragonfly, starfish), then pigs and chimpanzees suddenly seem quite similar to humans. Relative impressions of "close" and "far" are subjective and depend on context.
- Pigs and chimpanzees differ in chromosome counts. The opinion is often expressed that when two animals differ in this way, they cannot produce fertile hybrids. This rule is, however, only a generalization. While such differences do tend to have an adverse effect on the fertility of hybrid offspring, it is also true that many different types of crosses in which the parents differ in chromosome counts produce hybrids that capable themselves of producing offspring.
- There have been no systematic, scientific surveys of the crossability of mammals belonging to different taxonomic orders (a cross between pig and chimpanzee would be interordinal). Any firm opinion on such a point must therefore, necessarily, be prejudiced. In fact, certain fishes belonging to different orders have been successfully crossed.
- Ultimately, the interaction of gametes at the time of fertilization, and the subsequent interplay of genes (derived from two different types of parents) during the course of a hybrid’s development cannot be predicted by any known laws because the interaction is between a multitude of extremely complex chemical entities that each have an effect on others. It is for this reason that the degree of similarity perceived between two organisms is no sure indicator of their crossability.
- Another suggestive fact, probably known to the reader, is the frequent use of pigs in the surgical treatment of human beings. Pig heart valves are used to replace those of human coronary patients. Pig skin is used in the treatment of human burn victims. Serious efforts are now underway to transplant kidneys and other organs from pigs into human beings. Why are pigs suited for such purposes? Why not goats, dogs, or bears — animals that, in terms of taxonomic classification, are no more distantly related to human beings than pigs? (In subsequent sections, these issues are considered in detail.)
- God did not place pigs and humans in different taxonomic orders. Taxonomists did. A great deal of evidence (read a discussion of this topic) exists to suggest that taxonomists are, in no way, infallible. Our ideas concerning the proper categorization of animals are shaped by bias and tradition to such an extent that it would be rash to reject, solely on taxonomic grounds, the feasibility of such a cross.
- The general examination of the process of evolution as a whole (as presented elsewhere on this site) strongly suggests that most forms of life are of hybrid origin. Why should humans be any different?
- It might seem unlikely that a pig and a chimpanzee would chose to mate, but their behavior patterns and reproductive anatomy do, in fact, make them compatible (this topic is considered in detail in a subsequent section). It is, of course, a well-established fact that animals sometimes attempt to mate with individuals that are unlike themselves, even in a natural setting, and that many of these crosses successfully produce hybrid offspring.
- Accepted theory, which assumes that humans have been gradually shaped by natural selection for traits favorable to reproduction, does not begin to account for the relative infertility of human beings in comparison with nonhuman primates and other types of animals (see previous section). How would natural selection ever produce abnormal, dysfunctional spermatozoa? On the other hand, the idea that humans are descended from a hybrid cross — especially a relatively distant cross — provides a clear explanation for Homo's puzzling and persistent fertility problems.
- If we supposed standard theory to be correct, it would seem most peculiar that pigs and humans share features that distinguish human beings from chimpanzees, but that pigs and chimpanzees should not. Conventional theory (which assumes that pigs are equally as far removed from humans as from chimpanzees) says that pigs and chimpanzees would share about as many such traits as would pigs and humans. And yet, I have never been able to identify any such trait—despite assiduous investigation. The actual finding is that traits distinguishing chimpanzees from humans consistently link pigs with humans alone. It will be difficult to account in terms of natural selection for this fact. For each such feature, we will have to come up with a separate ad hoc argument, explaining how the feature has helped both pigs and humans to survive and reproduce. On the other hand, a single, simple assumption (that modern humans, or earlier hominids that gave rise to modern humans, arose from a cross between pig and chimpanzee) will account for all of these features at a single stroke.
| A reader's comment: "The theory overcomes the creationist's objection to gradualism and the evidence for pig ape hybridity has no stronger scientific competition. Open your mind and look at the facts. Consider how it might be true. Let go of your prejudices and misinformations. Not all hybrids are sterile. Examples of hybrid crosses are common in nature, including fertile ones. Admittedly transordinal crosses are unusual, but then we are extraordinary." |
For my own part, curiosity has carried me away from my old idea of reality. I no longer know what to believe. Is it possible that so many biologists might be wrong about the nature of human origins? Is it possible for a pig to hybridize with a chimpanzee? I have no way of knowing at present, but I have no logical or evidential basis for rejecting the idea. Before dismissing such a notion, I would want to be sure on some logical, evidentiary basis that I actually should dismiss it. The ramifications of any misconception on this point seem immense. As Huxley put it long ago, "The question of questions for mankind — the problem which underlies all others, and is more deeply interesting than any other —is the ascertainment of the place which Man occupies in nature."
Are we simply another type of primate, like the chimpanzee or the baboon? Or are we a complex melange, an alloy of two very distinct forms of life? These are questions that can only be resolved by examining the evidence. I invite the reader to consider these two possibilities as simple hypotheses, to consider the data coldly, and then to determine which of the two is more consistent with available evidence.
Are we simply another type of primate, like the chimpanzee or the baboon? Or are we a complex melange, an alloy of two very distinct forms of life? These are questions that can only be resolved by examining the evidence. I invite the reader to consider these two possibilities as simple hypotheses, to consider the data coldly, and then to determine which of the two is more consistent with available evidence.
| Then felt I like some watcher of the skies When a new planet swims into his ken. |
| — John Keats On First Looking into Chapman's Homer |
| Is hairless skin a trait seen only in modern domestic pigs (and not anciently)? A reader wrote in with the following question about hair:"While some domestic pigs are bred to be relatively hairless, all the wild pigs seem to be fully-furred. In fact, when domestic pigs go feral, they seem to immediately revert to a hairy form. If so, how could we have inherited the hairless trait from pigs?" I sent back this response: "When a pig escapes from a farm and starts living in the woods it does not suddenly become a hairy animal. It's descendants can, if they interbreed with hairy wild animals, but not otherwise. True, the Eurasian wild boar is hairy (though its hair is nowhere near as dense as that of a cow or sheep, say). But we do not know the history of the domestic pig. It's usually treated as conspecific with the Eurasian wild boar, but the two differ in chromosome counts (domestic 2n=38, and wild boar 2n=36). So it may be that they are not the same animal and that relatively hairless pigs similar to the domestic pig existed anciently. It may well be that the two have been treated as the same species merely because it has long been known that they can produce fertile offspring together. But these offspring may simply represent hybrids (this is one of many examples, by the way, of animals with differing chromosome counts producing fertile offspring together). The domestic pig has also hybridized with a variety of other types of pigs, but that does not imply that they are the same animal. For example, in addition to the wild boar, the domestic pig has also hybridized with the Babirusa,Babyrousa babyrussa (pictures); Bush Pig, Potamochoerus larvatus(pictures); Bearded Pig, Sus barbatus(pictures); Visayan Warty Pig, Sus cebifrons (pictures); Sulawesi Wild Boar, Sus celebensis (pictures); and probably Sus oliveri and Sus philippensis. So why assume that the domestic pig and wild boar are the "same" animal? Relatively naked animals similar to the domestic pig might have existed anciently. We don't really know what pigs looked like thousands of years ago, but a prehistoric painting in Altamira Cave in Spain shows a pig (pictures) that looks fairly naked to me (except for what looks like a beard and hair at the top of the head, neck, and shoulders)." |
Some of the most easily accessible evidence that can be used to evaluate the hybrid hypothesis is visible in the mirror. In this section, we will consider certain external features that link humans with pigs. Much of my research on pigs has centered on the ordinary pig (Sus scrofa). Of course, ordinary pig is really a catchall term for a variety of breeds. "There are currently some 87 breeds of domestic pigs in the world, most of them in Europe and North America," according to Pond and Houpt, and "another 225 or more groups of pigs not recognized as breeds but each having unique characteristics, appearance, or geographical location."1 However, the focus here will be on traits that are generally characteristic ofSus scrofa.
And now, let's look a little more closely at some human distinctions that, as it turns out, are characteristics of pigs as well. Traits that distinguish us from chimpanzees and other primates are the only ones that will be discussed, because traits that humans share with primates have no bearing on the question of whether humans are of hybrid origin. Under the hypothesis being considered, it would make no difference if humans are more similar to chimpanzees in most respects than to pigs. The interesting finding is that those features that do distinguish humans from chimpanzees and other primates can be consistently accounted for by reference to the pig.
This physical affinity of humans and pigs is easily observable in certain external features. This fact did not escape Thomas Mann, who once wrote "The pig with its little blue eyes, its eyelashes and its skin has more human qualities than any chimpanzee — think how often naked human beings remind us of swine."² Although I do not concur in Mann's assertion that pigs share more traits with humans than do chimpanzees, I do think pigs and humans share more than enough traits to suggest a relationship. For example, lightly pigmented eyes, in shades of blue, green, and tan, are never found in chimpanzees or orangutans.3 There is, apparently, only one known case of a gorilla with blue eyes.4 Light-colored eyes are also rare in other primates.5 Why, then, are they common in certain human populations? Where did this trait come from? One conceivable explanation is that it was inherited from blue-eyed pigs. Blue is a common eye coloration in swine (as are green, yellow, and tan). The dark pigment (melanin), found so consistently in the irises of nonhuman primates, is beneficial. It absorbsultraviolet light. To protect their eyes from these damaging rays, pigs depend on their narrowly slit, heavily lashed eyelids. Humans shield their eyes in a similar way, unlike the typical wide-eyed, sparsely lashed ape.
| In the gorilla, Schultz remarks that he "found a roof cartilage of less than 1 cm² and paper-thin alar cartilages, limited to the nasal center and not extending into the huge wings, which were mere pads of fat. In contrast to this, the prominent nose of man is far more extensively supported by cartilage, which closely determines its shape. While the nearly immobile nasal wings of apes consist of little more than skin and fat, the thin and mobile wings of human noses are extensively stiffened by cartilage to keep them from being sucked shut with every inhalation (495.9,52). | |
| While Schwartz's statement concerning the uniqueness of the human nose is generally correct, it must be said that certain Asian monkeys (Nasalis,Rhinopithecus) do have protrusive noses (235.4,29). | |
| Walker (588.4,1175) states that "this cartilaginous snout [of pigs], used for turning up surface soil, is strengthened by an unusual bone, the prenasal, situated below the tip of the nasal bones of the skull." Composed primarily of cartilage, this flexible prenasal "bone" finds its equivalent in the cartilaginous tip of the human septum. |
Neither is it clear how a protrusive cartilaginous nose might have aided early humans in their "savannah hunter lifestyle." As Morris remarks, "It is interesting to note that the protuberant, fleshy nose of our species is another unique feature that the anatomists cannot explain."6 This feature is neither characteristic of apes, nor even of other catarrhines.7 Obviously, pigs have a nose even more protuberant than our own. In a pig's snout, the nasal wings and septum are cartilaginous as ours are.8 In contrast, a chimpanzee's nose "is small, flat, and has no lateral cartilages" (Sonntag9). A cartilaginous nose is apparently a rare trait in mammals. Primatologist Jeffrey Schwartz goes so far as to say that "it is the enlarged nasal wing cartilage that makes the human nose what it is, and which distinguishes humans from all other animals."10The cartilaginous structure of the pig's snout is generally considered to be an "adaptation" for digging with the nose (rooting). Rooting is, apparently, a behavior pattern peculiar to pigs. Other animals dig with their feet.
A protruding nose is perhaps the most prominent difference between a human face and that of a chimpanzee, but discussions of human evolution rarely mention the nose, perhaps because its lack of utility precludes explanation in terms of adaptation. Instead, most analyses deal with the fleshless skull, where the protrusiveness of the human nose is a bit less obvious (but visible nonetheless). It is a peculiar omission, because useless (nonadaptive) traits are widely considered to be the best indicators of relationship. What is the evolutionary utility of our unique nasal structure? Is it functional? Or is it the genetic residue of an ancient hybrid cross?
| Specifically, Sonntag notes the lack of a philtrum in chimpanzees (533.6,371). | |
 |
Another feature to consider is the philtrum, the dent seen on the center of the human upper lip. Apes lack this typical human feature.11 It seems a useless structure from a survival standpoint. Why is it seen, then, the world over in Homo? In both human beings and pigs, during the early stages of development, the upper lip is cleft, though I have not been able to find any evidence of such a cleft in the embryos of any nonhuman primate. As development continues, this cleft usually closes in humans, but persists in pigs.12The human philtrum is a visible residue of this primordial split lip. In those human beings where this split never closes, the condition is known as cleft lip, a common birth defect. The frequent occurrence of cleft lip in humans is hard to explain if it is assumed that we are closely related only to primates. If the assumption, however, is that human beings are derived from a pig-chimpanzee cross, this finding becomes far more understandable.
Similar thinking explains the shortness of the human upper lip (distance between mouth opening and nostrils). Why has our upper lip become shorter and thicker in the course of evolution? All apes have upper lips much longer than those of humans,13but a pig's upper lip is so short that it is scarcely more than an appendage of the snout.14 Morris15 makes much of the fact that human lips are covered on their exterior surface by glabrous (i.e., absolutely hairless) mucous membrane:
Like the earlobes and the protruding nose, the lips of our species are a unique feature, not found elsewhere in the primates. Of course, all primates have lips, but not turned inside-out like ours. A chimpanzee can protrude and turn back its lips in an exaggerated pout, exposing as it does so the mucous membrane that normally lies concealed inside the mouth. But the lips are only briefly held in this posture before the animal reverts to its normal 'thin-lipped' face. We on the other hand, have permanently everted, rolled-back lips.
He goes on to suggest that our peculiar lips are the product of "sexual selection." But other explanations are conceivable: In describing the skin of pigs, Getty16 states that "there are no true glabrous surfaces other than the labial borders," which are composed of red mucous membrane.
| Some disagreement exists in the literature over the question whether earlobes are present in apes. Sonntag says they are not seen in the chimpanzee (533.8,86), but Schultz (495.65,146) claims they are sometimes found in the African apes and even in certain monkeys. |
In reference to human earlobes, Morris observes that "anatomists have often referred to them as meaningless appendages, or `useless fatty excrescences.' By some they are explained away as `remnants' of the time when we had big ears. But if we look to other primate species we find that they do not possess fleshy earlobes. It seems that, far from being a remnant, they are something new."17 Perhaps, however, they are really something old on a new face. Sisson describes the lower portion of a pig's ear as "strongly convex below, forming a prominence somewhat analogous to the lobule of the human ear."18
An additional feature of the human ear should be mentioned here, the Darwinian tubercle(see Darwin's illustration below). In his Descent of Man, Darwin comments on this feature sometimes found on the rim of human ears which he describes as "a little blunt point, projecting from the inwardly-folded margin, or helix … These points not only project inward, but often a little outward, so that they are visible when the head is viewed from directly in front or behind. They are variable in size and somewhat in position,
standing either a little higher or lower; and they sometimes occur in one ear and not on the other. Now the meaning of these projections is not, I think, doubtful, but it may be thought that they offer too trifling a character to be worth notice. This thought, however, is as false as it is natural. Every character, however slight, must be the result of some definite cause; and if it occurs in many individuals deserves consideration. The helix obviously consists of the extreme margin of the ear folded inward; and this folding appears to be in some manner connected with the whole external ear, being permanently pressed backward. In many monkeys, which do not stand high in the order, as baboons and some species of macacus, the upper portion of the ear is slightly pointed, and the margin is not at all folded inward, a slight point would necessarily project inward and probably a little outward. This could actually be observed in a specimen of the Ateles beelzebuth in the Zoological Gardens; and we may safely conclude that it is a similar structure — a vestige of formerly-pointed ears — which occasionally reappears in man.19
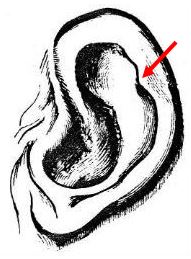 |
Darwinian tubercle (Darwin, 1871) |
Primatologist Adolph Schultz (1973), however, flatly contradicts Darwin, saying that "clearly pointed ears, commonly called `satyr ears,' are among monkeys typical for only macaques and baboons and do not occur in any hominoids [great apes], not even in the early stages of development. There is no justification, therefore, to interpret the occasional `Darwinian tubercles' on human ears as an atavistic manifestation of ancestral pointed ears."
| According to Schummer et al. (503.3,497), "The eyebrows [of the domestic pig] are formed by 2 to 3 rows of prominent tactile hairs formed at the base of the upper eyelid; there are more than 40 in all and they are up to 8 cm long. They form into bundles, especially at the medial angle of the eye." |
Swine have prominent eyebrow hair. On the brows of the chimpanzee fetus it is possible to discern a region of light-colored bumps following a pattern similar to that of the human eyebrow. Adult apes, however, have no eyebrow hair.21 On their eyelids, pigs have luxuriant eyelashes, thicker even than those of human beings. In many pigs these cilia, as anatomists term them, are so thick that the animal seems to be wearing false eyelashes. But apes scarcely have eyelashes at all, despite the apparent survival value of this feature. Also, pongids have prominent brow ridges while pigs and most humans do not. If we choose to explain the development of human eyelashes and eyebrows in terms of natural selection, we must wonder why apes, which have existed at least as long as any hominid, have failed to acquire them. Perhaps their heavy brow ridges sufficiently protected their eyes, but if such is the case, why did not brow ridges also suffice for Homo? What was the pressing need that caused Homo to substitute tufts of hair for ridges of bone?
Dermal Characteristics
That humans lack the hair cover of nonhuman primates is an accepted fact. "It is this single factor that constitutes the chief difference between human skin and the skin of other mammals" (Montagna22). Some writers say that the hair coat of a chimpanzee is "sparse." But if "sparse" describes chimpanzee pelage, then "naked" accurately describes the skin of human beings. Any human who even approached the hairiness of other primates would be considered abnormal. Pigs, however, are a different case. Many domestic pig breeds have skin just as naked as human skin.
| Schultz (495.07,Plate 1) pictures a 185-day-old chimpanzee fetus that is virtually hairless except for a thick patch atop its head (in the same region it is seen in human beings). It also has eyebrow hair arranged in the same pattern as do humans. |
Other primates do not have the long mane of hair that tops the head of an unshorn human, nor do they have beards. Haltenorth notes that in some varieties of Sus scrofa, manes are found on the neck and back ("Näcken-/Rückenmähne"), beards on the cheeks ("Wangenbart"), and shocks of hair on the forehead and atop the head ("Stirn-/scheitelschopf"). He also says that the last of these three traits is found, among pigs, in Sus alone.25 A prehistoric painting of a pig found in Altamira Cave in northern Spain depicts an animal with a beard and thick hair atop its head. Sus barbatus, an extant pig native to southeast Asia (which forms fertile hybrids of both sexes in crosses with S. scrofa) has little hair on its body, but does have a very thick and bushy beard.26
Panniculus adiposus. In an article on the evolution of human skin, renowned cutaneouscomparative anatomist William Montagna notes that, "Together with the loss of a furry cover, human skin acquired a hypodermal fatty layer (panniculus adiposus) which is considerably thicker than that found in other primates, or mammals for that matter. This is not to say that only man has a fat skin, but a thick fatty layer is as characteristic an attribute of human skin as it is of pig skin."27 Similarly, Dyce et al. (160.1,742) note that there is a "well developed fat deposit present almost everywhere in the subcutis." Primatologist F. W. Jones also noted this fat layer:
"The peculiar relation of the skin to the underlying superficial fascia is a very real distinction [of human beings], familiar to everyone who has repeatedly skinned both human subjects and any other members of the primates. The bed of subcutaneous fat adherent to the skin, so conspicuous in man, is possibly related to his apparent hair reduction; though it is difficult to see why, if no other factor is invoked, there should be such a basal difference between man and the chimpanzee."28
Panniculus carnosus. "Another particularity of human skin is its general lack, or loss, of the cutaneous skeletal muscle layer (panniculus carnosus) found throughout the skin of most other mammals. Remnants of a panniculus carnosus in human skin are found at the front of the neck in the apron-like, thin platysma muscle … All other primates, even the great apes, have a panniculus carnosus over much of the body" (Montagna29). As in humans, the cutaneous musculature of pigs is well developed in the neck (platysma muscle) and face, but sparse or nonexistent elsewhere.30
In animals having a panniculus carnosus, the skin receives its blood supply from direct cutaneous arteries (large superficial vessels running parallel to the skin surface in the cutaneous muscle sheath). But when no panniculus carnosus is present, arteries feeding the skin rise up like little trees from deep within the body. Arteries of this latter type are calledmusculocutaneous. These two forms of dermal circulation are depicted in the illustration below. Both pig skin and human skin are supplied by musculocutaneous arteries.31 As Daniels and Williams observed in a 1973 article on skin flap transfer, "Most experimental animals do not have a vascular supply to the skin similar to that of man. The pig's cutaneous vascular supply has been demonstrated anatomically and surgically to be more comparable than most to that of man … As in man, the pig's skin is supplied by ubiquitous musculocutaneous arteries and by a few direct cutaneous arteries."32 This observation has been confirmed by other authors: "Except for pigs, whose cutaneous vasculature resembles that of man, loose-skinned mammals are vascularized by direct cutaneous arteries" (Montagna and Parakkal33). Therefore, in this respect, human skin is more similar to pig skin than to that of nonhuman primates: "Actually, the vascularity of the skin of most nonhuman primates is essentially similar to that of other furred animals" (Montagna34). In particular, Baccaredda-Boy,35as well as Moretti and Farris,36 found that the skin of chimpanzees differs from that of human beings in having numerous large, superficial vessels (i.e., direct cutaneous arteries).
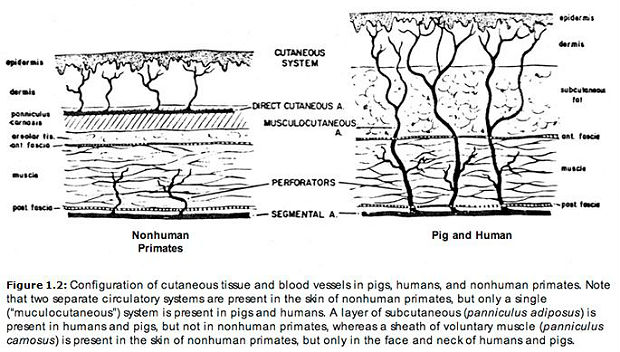 |
| In the paragraph at left, the calculations for the pig capillaryseparation interval were based on Young and Hopewell's data (605.4, Fig. 1 and Table 2). In the chimpanzee, the epidermis is richly vascularized only beneath the friction surfaces (palms and soles), not beneath the hairy-skin regions. Thus, regarding the chimpanzee, Montagna (365.5,191) states: "Where the epidermis is flat [i.e., hairy-skin regions], capillary loops are ill-defined … Capillary loops are deepest and most complicated underneath the epidermis of the friction surfaces. |
Human skin also stands apart from that of other primates — and from that of most other mammals for that matter — with respect to the quantity of blood that can be circulated through it.37 A certain amount of blood is needed just to feed the skin. This is the amount it receives in most animals. In humans, however, the maximum blood flow can be more than a hundred times greater than this minimum.38 Fed by temperature-sensitive musculocutaneous arteries, the densely spaced cutaneous capillaries of human beings play an essential thermoregulatory role.39 When the body begins to overheat, large quantities of warm blood can be rapidly cooled in these capillaries via sweat evaporation. One measure of cutaneous vascular density is the capillary loop separation interval. In human beings, the typical distance between capillaries ranges from 50 to 100 microns.40 In porcine flank skin, this figure is reduced to only about 20 microns, a separation interval so small as to be almost incredible. When white pigs are exposed to high temperatures, the skin flushes pink with blood (even in the absence of sunlight) as it does in light-skinned human beings under similar conditions.41
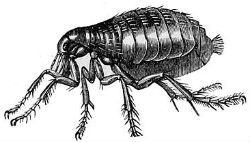 | |
Human flea, Pulex irritans |
Fleas. Perhaps this difference between our cutaneous vasculature and that of our primate kin accounts for another human distinction: "Ironically," writes Nicole Duplaix, "man is unique among the primates in having fleas."42 More than 2,400 distinct types of fleas have been treated as species or subspecies.43 Parasites are usually rather specific in their choice of host. Fewer than twenty of these 2,400 types will readily bite human beings.44 Foremost among those that feed on Homo sapiens is the human flea, Pulex irritans, but we are not the only suitable hosts for this species. According to Bennett, "Pulex irritans, the human flea, breeds freely in hog-house litter and may become a serious pest of swine."45
| Newton's law of cooling states that the rate at which heat flows out of a warm body into a cooler surrounding medium is proportional to the difference between the temperature at the body's surface and the temperature of the surrounding medium. |
The panniculus adiposus replaces hair as an insulating layer in human beings and pigs. According to Beckett (63.8,2),
The pig increases or decreases the amount of heat lost … by varying the blood flow in the [skin's] capillary bed … If all blood flow to the outer body parts were stopped, the thermal resistance between the body cavity or muscle tissue and skin surface would approximately equal the resistance of the fat layer plus the resistance of the hair and skin. To the extent that a pig is able to direct a sizable flow of blood through the skin and region just below the skin, the fat layer is by-passed and thermal resistance is at a minimum.
In the figure above, notice that the musculocutaneous arteries pass through the cutaneous fat.
This perforated fat layer constitutes an insulating mechanism that can respond quickly to ambient temperature, a characteristic that hair lacks. Dilation of the musculocutaneous arteries in response to heat increases blood flow to the skin. This increase in circulation can raise skin surface temperature to a level almost as high as that within the body, thus increasing the rate at which heat is lost to the environment.b In cool environments, constriction of these arteries reduces skin temperature and, consequently, the rate at which body heat is lost to the atmosphere because the fat layer can then serve as an insulating blanket.
| Obviously, furred animals cannot remove their coats when it's hot — they shed. But shedding is a process that takes weeks, not minutes. It is a seasonal adjustment, not the moment-to-moment adjustment seen in human beings and pigs. |
Possession of a panniculus adiposus allows adjustment to changes in ambient temperature on a moment-to-moment basis — a clear advantage in the temperate zones where much of the human race has made its home, because these regions are much more subject to sudden, extreme shifts in temperature than those close to the equator. Nonhuman primates and other furred animals do not have the option of adjusting their skin temperature. Because their skin is not insulated from the rest of the body by a layer of fat, its temperature must remain near that of the flesh beneath it.
Pig skin is separated from the inner body by a thick fat layer, and it can cool to an extreme degree. Fat, not hair, is the primary insulating barrier.47 Alaskan swine can withstand sub-zero temperatures by cooling their skin to as little as 9˚ C (at an ambient temperature of -10˚ C) without suffering tissue damage.48 Acclimatized human beings, too, can reduce skin temperature to about 10˚ C without injury.49 This mode of insulation is completely different from that of nonhuman primates, more like that seen in certain aquatic mammals (e.g., seal, walrus). With the exception of the pig, it seems that no other land animal has this form of insulation.
More than a naked ape, Homo is a variably insulated naked ape. In hot environments human beings (and pigs) can the increase circulation of warm blood to the skin and raise temperatures almost to the level of body core temperature, thus maximizing heat loss to the surrounding air. If weather turns cold, they can restrict cutaneous circulation, cooling the skin to such a degree that heat loss is significantly reduced. This ability is especially apparent in fat50or acclimatized individuals.51 Although a cultural advance, the invention of clothing, made it possible for Homo to inhabit cool regions formerly off-limits to primates, a biological advance, in the form of a new insulation system, has increased the human ability to withstand the sudden temperature variations found in those regions.
| If skin has any hair whatsoever (scalp, forearm, belly) dermatologists refer to it as "hairy skin." Hairy skin in humans, then, is the skin covering most of the body, the general body surface. Other regions, that are absolutely hairless (lips, palms, soles) are called "glabrous." |
Besides being a good insulator, human skin is surprisingly thick. "The epidermis over our general body surface ["hairy skin" see note at right] is substantially thicker than that of other primates: the horny layer [stratum corneum] can be peeled off intact as a diaphanous but tough membrane that can be used for experimental purposes … The epidermis in the hairy skin on nonhuman primates, mostly like that of any other furred mammal, is relatively thin, with a relatively thin horny layer" (Montagna52). Pigs, though, have a thick epidermis and stratum corneum, thicker even than that of human beings.53
| Another quotation from Montagna (360.3,13): "Elastic fibers are numerous everywhere [in pig skin]. In the papillary layer delicate fibers branch toward the epidermis as they do in the skin of man." | |
| In the case of the chimpanzee's dermis, Montagna and Yun state that, "Elastic fibers, nowhere numerous, are concentrated in the papillary body and in the deep portion of the reticularis dermis" (365.5,191). |
The elasticity of our skin is also unusual. "Whereas the skin of the great apes and that of some of the simian primates have variable amounts of elastic fibers, in no animals, regardless of sex, age, or locality have we found the abundance of elastic tissue characteristic of human skin" (Montagna54). This finding comes from the same author who, in an earlier article comparing human skin with that of pigs, observed that "one of the most striking resemblances between these two skins [pig and human] is the large content of elastic tissue in the dermis."55
He also remarks that "the surface of both skins [human and porcine] is grooved by intersecting lines which form characteristic geometric patterns."56 In a separate paper on the evolution of human skin he provides a little more detail:
The outer surface of human skin is crisscrossed, almost everywhere, by fine intersecting congenital lines … (You can confirm the presence of these lines by looking at the back of your hands). This characteristic is not limited to human skin; creases are also found on the skin of pachyderms, walruses, and, to a lesser extent, pigs. With the exception of occasional, shallow creases, the surface of the hairy skin of nonhuman primates is smooth.57
The presence of these lines in both pigs and humans is not easily explained in terms of natural selection since they have no known function.58
On the underside of our "hairy skin" (general body surface), where the epidermis meets the dermis, is a different patterning not corresponding in its configuration to the outside patterning described in the preceding paragraph. A similar, though coarser, pattern is also characteristic of the epidermal-dermal junction in pigs. Montagna, however, notes that "in split-skin preparations where the epidermis is neatly removed from the dermis, the epidermis of heavily haired animals is flat.59 Even in monkeys and apes, epidermal grooves are found only around the attachment of the ducts of glands and pilary canals." We can account for a finer patterning in humans than in pigs by the fact that a fine mesh is intermediate between the coarse patterning of pig skin and the smooth undersurface of nonhuman primate skin.
| Note: A section discussing sweating in humans, pigs, and chimpanzees, which formerly appeared here, has been moved. Sorry for any confusion or inconvenience this may have caused! Jump to the new location >> |
So, in the pig, we have a sparsely haired animal with a fatty, stretchy skin supplied by musculocutaneous arteries. The surface of the hairy skin is marked by congenital lines similar to those seen in human beings, and the patterning of the epidermal-dermal junction is also quite similar in the hairy skin regions. Under the hypothesis that we are considering, it makes little difference that pig skin differs from human skin in other ways. The essential point is that, in those cases in which our skin is peculiar for a primate, an explanation for each such anomaly can be found in the skin of pigs.
The Savanna Hunter
| A mature pig has about 500,000 large sweat glands distributed over its entire body (503.3,497; 506.5,316). Nevertheless, it is often asserted in the literature that pigs do not sweat. This assumption can be traced to studies by Ingram and by Mount, who studied perspiration rates in immature animals, usually sedentary piglets (247.03; 247.1; 389.7; 390.1; 390.2; 390.3; 390.5). Studies evaluating pig sweating have concentrated on young pigs because they are of greater commercial interest. Immature animals are no more appropriate for determining the evaporative qualities of a boar or a sow than a toddler would be for revealing traits of an adult human—Children sweat much less than do adults (584.4,577). Small animals have a tendency to hypothermia (because their surface area is large in proportion to their size), not hyperthermia, and have little tendency to sweat (390.8,182). Perspiration in pigs is often overlooked because these animals are, apparently, more efficient sweaters than are humans. Their sweat glands seem to be better attuned to thermoregulatory needs (they produce no more sweat than what is necessary to cool cutaneous blood by evaporation). Very little sweat is lost to runoff because sweat rarely builds up on the skin. But observed rates of sweating in mature pigs are approximately comparable to those of humans. Beckett (63.4) found that a 350 lb. sow at rest lost approximately 95 g/m² in sweat per hour at a dry bulb temperature of 98E F and wet-bulb temperature of 81). At a much higher temperature (122EF dry bulb and 79EF wet bulb), Myhre and Robinson found that 70 kg men at rest lost moisture (sweat + respiration) at a rate 250 g/m<² per hour (398.7,Table 3). Even in smaller pigs (198 lb. gilts), skin moisture loss is important (387.8,Table 1), ranging from one-third to two-thirds of total moisture loss (lung + skin). The claim that pigs need a wallow when living in hot climates (because they supposedly do not sweat) is also encountered. But Heitman and Hughes exposed hogs without access to a wallow to high temperatures (100E F; relative humidity 35%) for a week without any fatalities—conditions where the only avenue for heat dissipation is evaporative cooling (232.5,176). |
Pigs sweat when they are hot. "The apocrine [i.e., sweat] glands of the horse and pig secrete profusely during violent exercise and stress" (Montagna160). This sweating serves a thermoregulatory function in pigs just as it does in human beings.161 The hairy skin sweat glands of nonhuman primates, however, do not respond to thermal stimulation. The failure of nonhuman primates to sweat puzzled Montagna: "One might surmise," he writes,
that, like man, these animals sweat in response to heat stimulation. However, with singular exceptions, if the glands secrete at all, the amount is so small that it cannot be recorded. Sometimes animals show beads of sweat on the facial disc when under deep anesthesia, but our efforts to induce thermal sweating have failed. We have also largely failed to induce sweating with sudorific drugs, either cholinomimetic or adrenomimetic. In the chimpanzee, very few, small sweat drops were recorded even after the administration of shockingly large doses of these drugs.162
In contrast, even a small dose of acetylcholine or adrenaline elicits sweating in pigs. Even the immature pigs studied by Ingram (247.1,95) responded to adrenalin.
The notion that nakedness has somehow enhanced sweat evaporation in humans is widely received. Supposedly, our sparse pelage allowed our ancestors to cool their skin more rapidly than hairy animals in hot, dry environments, or somehow improved their ability to dissipate metabolic heat while rushing about the savannah in pursuit of prey.
Russell Newman, however, points out that our lack of reflective hair actually increases solar heat load and the need to sweat.164 To substantiate this claim, he cites a study by Berman showing that cattle exposed to the sun sweat more after their hair is removed.165 Similarly, panting increases in shorn sheep.166
Clothing, which replaces hair as a radiation barrier in human beings, has much the same effect on human perspiration. Human beings subjected to solar heat loads sweat more when naked than when wearing light clothing under otherwise identical circumstances. In a study of the effects of clothing on sweat, Adolph167concluded that "the nude man can save easily as much body water by putting on a shirt and trousers as can the clothed man by finding good shade." Moreover, body hair does not reduce convective heat loss "and has nothing to do with radiation of long-wave infra-red heat to cooler objects," says Newman.168 He therefore asserts that naked skin,
is a marked disadvantage under high radiant heat loads rather than the other way around, and that man's specialization for and great dependence on thermal sweating stems from his increased heat load in the sun.169
| My very crude experiment: I took two beakers and lined the bottom of one with a circle of rabbit fur. I then placed water, drop by drop, in equal amounts in both beakers. I continued the experiment for several days, always keeping the fur damp. Day after day, the water level rose in the beaker without fur. But no water buildup at all occurred in the other beaker. | |
| Claims that naked skin confers an evaporative advantage can, for the most part, be traced to a single sentence: Mount (390.8,42) seems only to be expressing an opinion in saying that "In a bare-skinned animal, like pig or man, the evaporation of water from the body surface takes up most of the heat required for the process from the body itself, and so constitutes an efficient cooling system." Nevertheless, many later authors cite this statement in substantiation of the claim that bare skin enhances sweat evaporation. I have not been able to locate any actual data (in the works of Mount or any other author) demonstrating this assertion. |
If increased radiant heat loads caused early humans to depend more on sweat for cooling, why has hair loss, which increases those loads, progressed to the degree that it has in Homo? Under the assumption that humans first evolved on the arid, sun-drenched savannah, it is difficult, in terms of survival efficiency, to account for a reduction in hair density that would result in increased rates of water consumption. Newman points out that there is no evidence that hair interferes with sweat evaporation. Actually, I myself performed a crude experiment, the results of which indicate that hair actually accelerates the evaporation of sweat. This finding is surprising in light of evolutionary theorists' frequent claims to the contrary. But with a little consideration, one realizes that a hair coat is not a vapor barrier. Fur's ability to "breathe" has always distinguished it from less desirable insulators that slow heat loss but don't "wick away" moisture from the skin. Why should hair not only allow, but even enhance, evaporation rates? There are at least two reasons. First, wet hair presents a more irregular surface to the surrounding atmosphere than does hairless skin, augmenting the surface area available for evaporation. Second, hair allows uniform dispersion of sweat by capillary action, preventing the formation of the individual droplets seen on naked skin. When such droplets form, the skin lying between them does not serve as an evaporative surface and the vaporization rate is reduced.
| At 40E C (the approximate temperature of the body surface under hyperthermic conditions) the latent heat of vaporization of water is 2406 J g-1 = 575 cal g-1. The evaporation of just a half-cup of sweat is sufficient to reduce the temperature of a 150 pounds of water by an entire centigrade degree. Evaporation is essential to heat absorption. Runoff merely removes fluid from the body without cooling it (When you pour out a cup from an urn of hot coffee, the temperature of the remaining coffee stays the same.). | |
| This rate (25% of sweat lost to run-off) is for men engaged in intense physical exercise at high temperatures (running without a shirt in the desert) (15.4). Runoff loss can thus be significant even in the desert, let alone on the more humid savannah. Adolph (15.4,93-94), for example, studied sweating in a man exercising strenuously in the desert (relative humidity: 32 percent; estimated wind speed, 10 m.p.h.; the man wore no shirt) and found that "his rate of evaporative loss was 1,300 grams per hour. His measured rate of weight loss, however, was 1690 grams per hour, exclusive of water which accumulated in his trousers." These figures indicate that 23 percent of the sweat went to runoff — even if we ignore the fact some of the water absorbed by the trousers would have run off of a naked human being. On the African savannah, the humidity and solar heat loads would be even higher because the savannah lies closer to the equator than the southwestern American desert where Adolph performed his experiments. On the savannah, then, a larger percentage of sweat would go to runoff (due to lower evaporative rates at the higher humidity) and, at the same time, a larger amount of sweat evaporation would be required to counteract the higher savannah heat loads. This waste of body fluids seems peculiar in a creature that is supposed to be the product of adaptation to a life of strenuous diurnal hunting on the open savannah. |
As the amount of sweat on the skin increases, the individual drops do merge to form a continuous sheet of water. But when a large amount of sweat is present on naked skin, another type of inefficiency sets in — runoff.
More sweat runs off hairless skin without evaporating. The coat of a hairy animal acts as a sponge, retaining sweat in position until it can evaporate. Perspiration dripping off the body has no cooling effect, because no heat is absorbed by runoff. In contrast, evaporating sweat absorbs a large amount of heat.1a But Adolph's research indicates that about a quarter of human sweat can be lost to runoff, even under near optimal evaporative conditions.1b A reflective hair coat, then, has three advantages: (1) lower solar heat loads; (2) increased rate of evaporation; (3) less sweat wasted on runoff. It is therefore difficult to understand how naked skin can be interpreted as an "adaptation" beneficial to a savannah hunter.
Of course, the "savannah hunter" hypothesis is just one of many theories. Hair loss in Homo has been the object of much speculation (for a survey of such theories, see 165.1). Besides those who say we lost our hair on the savannah170and/or because we were hunters,171there are others who suggest we may have lost it in the forest,172 or even in the sea.173 Some authors suggest that nakedness made us sexually enticing174or that hairlessness became thermally advantageous when we started wearing clothes.175
Even if we wished to assume that humans did at one time have a hair coat (there is absolutely no evidence that such was the case), these theories would not explain the advantage of a sparse coat of hair. The hunting hypothesis is untenable because nonhuman terrestrial predators all have thick hair coats. A similar objection can be raised to the sexual enticement scenario. Why haven't all mammals lost their hair if nakedness is enticing? The aquatic proposal is also dubious, most small (human-sized or smaller) aquatic or semi-aquatic mammals do have hair coats.176
The results of my evaporation experiment make it difficult for me to accept Mount's opinion that naked skin evaporates sweat faster than hairy skin.177 For the same reason, I doubt Wheeler's suggestion that the acquisition of erect posture by hominids "was probably the essential pre-adaptation which made it possible for them to shed body hair and develop extensive evaporative surfaces."178Also dubious is Kushlan's "vestiary hypothesis," because it proposes that the invention of clothing left Homo free to lose his body hair and thus obtain "the most efficient cooling system of any mammal."179 As we have seen, naked skin provides no particular evaporative advantage.
Because nakedness is a handicap on the savannah, Newman concludes, it is unlikely that human ancestors lost their hair after leaving the forest: "If one had to select times when progressive denudation was not a distinct environmental disadvantage, the choices would be between
a very early period when our ancestors were primarily forest dwellers or a very recent period when primitive clothing could provide the same protection against either solar heat or cold. The primary difficulty in arguing for the recent loss of body hair is that there seems to be no single and powerful environmental driving force other than recurrent cold that is obvious after the Pliocene epoch. Furthermore the developing complexity and efficiency of even primitive man's technology would have decreased the probability of a straightforward biological adaption … The obvious time and place where progressive denudation would have been least disadvantageous is the ancient forest habitat. Radiant energy does penetrate the forest canopy in limited amounts, [but] that portion of the spectrum which is primarily transmitted through the vegetation, the near infrared wavelengths of 0.75 to 0.93 microns, is exactly the energy best reflected by human skin (Gates, 1968180). 181
| In desert environments human beings can lose as much as 12 liters in sweat per day (390.3,162). Since the African savannahs lie closer to the equator than do most deserts, sweat rates there should be at least as high — if not higher. |
Note, however, that Newman does not explain why our ancestors lost their hair in the same environment (forest) where apes did not. If humans came into being via hybridization between pigs and chimpanzees, their genesis would almost surely have occurred in the forest. Chimpanzees live in forests. On the basis of its high rate of water consumption, Yang concluded the pig, too, is functionally a forest animal.182 Human beings need more water than almost any other animal.183
Indeed, it seems incredible that a hominid would spend any more time than necessary away from the forest. Although the savannahs of Africa were teeming with game, they were also swarming with ferocious predators. When a human being is chased by a lion, the first impulse is to find a tree. Consider the picture painted by current evolutionary theory: the noble savannah hunter, naked to the brazen sun, boldly erect on an arid and treeless plain, in indefatigable pursuit of a wary and dangerous prey, indifferent to the attack of rapacious carnivores. Certainly this description has dramatic appeal. It's like a Tarzan story. But is it plausible?
| The Lord will give grace and glory. No good thing will he withhold from them that walk uprightly. |
| —Psalms, 84:11 |
Plato's minimal definition of a human being as a "featherless biped" exploits the fact that it is unusual for a mammal to use only two feet in the course of normal locomotion. Since we're mammals, it's easy enough to understand the lack of feathers. Why, though, do we go about on two feet? Human bipedality has long been a subject of controversy. How long have human beings stood erect? How long did the transition take from quadrupedal locomotion to bipedality? What factors caused the change? Why have other primates not done the same?
Following in Darwin’s footsteps, a wide variety of authors have asserted that human beings gradually developed the ability to walk on two feet in response to selective pressures demanding that two hands be free to manipulate tools. In his book, The Ascent of Man,Darwin stated this view succinctly: "If it be an advantage to man to have his hands and arms free and to stand firmly on his feet, of which there can be little doubt from his pre-eminent success in
the battle for life, then I can see no reason why it should not have been more advantageous to the progenitors of man to have become more and more erect or bipedal. The hands and arms could hardly have become perfect enough to have manufactured weapons, or to have hurled stones and spears with true aim, as long as they were habitually used for supporting the whole weight of the body … or so long as they were especially fitted for climbing trees.1
This explanation is not without its flaws. For one thing, should we conclude on the basis of our supposedly “pre-eminent success in the battle for life” that every human trait is superior? Isn’t this line of reasoning a bit vague and self-indulgent? Are our hands really in any way perfect—or do we just see ourselves that way? Isn’t it possible to “manufacture weapons” while sitting down?
And then, there is the presumption that we became “more and more erect or bipedal.” Fossil evidence does not confirm this gradual transition. Apparently, even very early hominids were fully bipedal. Thus, Lovejoy observes, that "for a number of years and throughout much of the literature there has been an a priori assumption that australopithecine locomotion and postcranial morphology were 'intermediate' between quadrupedalism and the bipedalism of modern man. There is no basis
for this assumption...in terms of the lower limb skeleton of Australopithecus. It is often claimed, principally on the basis of this a priori assumption, that morphological features shared by both modern man and Australopithecus do not necessarily indicate similar gait patterns. Although this might be true in terms of a single feature, it is demonstrably not true when the whole mechanical pattern is considered...the only significant difference between the total biomechanical patterns of Australopithecusand H. sapiens is one that indicates that Australopithecus was at a slight advantage compared with modern man (femoral head pressure [i.e., pressure exerted by the weight of the body on the hip joint]).<²
| Pig tracks were also preserved at Laetoli (357.3,262a). |
Fossil footprints preserved in volcanic ash at Laetoli, Tanzania, indicate that hominids were fully bipedal at a very early date (3.7 million years ago).3 Similarly, Straus and Cave concluded that the posture of Neanderthals was not significantly different from that of modern humans.4 Homo erectus was also fully upright and bipedal.5 This lack of confirmation from the fossil record leaves gradualistic explanations of bipedalism standing on shaky ground.
Even on a hypothetical level, the idea that early humans "gradually" attained erect posture is implausible. One must either go on all fours or stand erect. No feasible intermediate posture exists. Hollywood portrays cave men as slumped over, arms hanging down. Maintaining such a position for any length of time would put an extreme strain on the muscles of the lower back.
Millions of years of slouching, then, would surely have produced more than a few backaches. In fact, it seems ridiculous to suggest that hominids went about day in, day out, partially erect. The physical strain would be too great, even for us with our supposedly better-balanced bodies.
Gradualistic thought forces the conclusion that early "human beings" spent a portion of their time in the quadrupedal position, but spent a gradually increasing portion of time erect as evolution progressed. Why would there be such a trend? Why have we developed the ability to stand all day on two feet?
| These "free" hands seem not to have been taken advantage of for more than a million years: The earliest known stone tools date from 2.6 million years ago (556.6,236), whereas indisputable evidence (Laetoli footprints) indicate that hominids were fully bipedal 3.7 million years ago (104.5; 293.8). | |
| This notion that free hands and intelligence are connected did not originate with Darwin, although he did espouse and popularize it. Perhaps, the earliest thinker to propose it was Anaxagorus who claimed that humans became intelligent by using the hands for manipulation rather than movement. Aristotle thought the opposite (i.e., that humans used their hands because they had become intelligent). | |
| Merfield (337.5,51) describes a favorite chimpanzee once on display at the zoological gardens in London. She was an inveterate smoker. "You could hand her a box of matches or a lighter, and she would not only use them properly, but could always be trusted to hand them back when she was finished with them." |
The many physical distinctions making a bipedal form of locomotion possible (even necessary, for efficiency) in humans would require many genetic alterations. Anyone who wishes to account for the spread of these mutations in terms of gradual evolution must show how bipedality increased our ability to survive and reproduce. Yet, a comparison of human and simian modes of locomotion, suggests bipedality does not appear to be any great boon.
Supposedly, "the freeing of the hands" made tool manipulation possible. The need for such manipulation, in turn, is said to have necessitated enlargement of the brain.
But why must a tool maker or a tool user stand erect for long periods of time? The hand, not the spine, seems to be the essential element in most manipulative processes. Few such activities would require anything more than the facultative bipedality of an ape. A chimp's hands would serve as well as ours in fashioning a spear, bow, or axe — they might even serve better: a chimpanzee has four hands. Human beings commonly sit down to work on such projects, having no need to stand. I can easily picture chimpanzees doing the same — chipping away at an arrowhead or heating spear tips in a fire. Studies of these animals have documented that their hands, too, are capable of performing subtle tasks such as decanting a glass of wine6 or even threading a needle.7 Surely, their using rocks and sticks to crack nuts8 is not so different from the way our forebears would have used hand axes.
Kortlandt has shown that chimps are capable of using weapons when they choose to do so.9In his experiments, he presented various objects to wild chimpanzees and recorded their reactions. In one test, he placed a stuffed leopard on the ground near a troop of chimpanzees. The "leopard" clutched a facsimile baby chimp in its paws, and a concealed tape recorder emitted baby cries. Presented with this phenomenon, the apes attacked, using large broken branches as clubs. Kortlandt says that the blows were of such force that, had the leopard been real, it would surely have been killed. Apparently we are not the only ones with "free" hands.
Jane Goodall has documented "aimed rock throwing" behavior in free-living chimpanzees.10 If they can carry clubs and throw rocks, then chimpanzees certainly have the anatomical wherewithal to carry and throw a spear. Physically, chimps may be better equipped for throwing than we are. Their arms are far stronger than those of human beings (about four times as strong, according to van Lawick).11 Our ancestors invented the spear thrower, a hooked stick that, in effect, lengthens the arm, increasing the force of the throw. The arms of chimpanzees are already longer and stronger than those of humans.
If they can carry clubs, apes should also be physically capable of stalking prey with a spear. Human hunters do not stand erect when their quarry is nearby. Rather, they crouch, or even crawl, and approach their prey from downwind, taking advantage of available cover. Only at the last moment when the prey is in range do they spring up and throw the spear. Chimpanzees are quite capable of leaping and throwing an object simultaneously.12
For all of these reasons, then, it is at least questionable whether bipedality has enhanced our ability to survive and reproduce. A gradualist would object that, even if we do not understand the selective pressures involved, such pressures must, nevertheless, have existed, and that humans otherwise would never have made the transition to erect posture. But slow selection of minute mutations is not the sole conceivable mechanism that can account for human bipedality.
An Analysis of Human Bipedality
If we listed all the features contributing to our upright mode of locomotion, we would find some of those features in the chimpanzee. Nevertheless, even though chimpanzees do walk on two feet from time to time, such is not their normal mode of progression. They lack certain characteristics that make moving around on the hind limbs not only convenient for human beings, but really, under most circumstances, the only practical way of getting around. But what if pigs possessed all of those features relevant to bipedality that apes lack. Wouldn't it then be easy to understand why a pig-ape hybrid might walk on two feet?
All the human distinctions listed in the remainder of this section were first identified by other writers; I've merely gathered them together. If a scholar somewhere has claimed that a certain characteristic distinguishes human beings from chimpanzees and that that feature contributes to bipedality, then — if I have encountered the claim — I at least mention it. I exclude only those features that relate to the skull; cranial features are discussed in the next section. (It will also be convenient in this section to discuss a few skeletal distinctions of human beings not directly relating to bipedality.)
In the literature, most features said to contribute to human bipedality are located in the spine and lower extremities. For example, our gluteal muscles, large in comparison to those of other primates13, enhance our ability to hold our torso erect. Ardrey observes that
| Sonntag notes the small size of the chimpanzee gluteal muscles in comparison with those of humans (533.6,356) and that they are small, also, in the gibbons and Old World monkeys (533.8,55,65). Duckworth (158.3,179) observes that the musculature of the upper limb is almost exactly as heavy as that of the lower limb in apes but that in humans the leg muscles are three times as heavy as those of the arms. Although I have not been able to obtain exact data on swine relating to this proportion, a cursory examination of any pig will reveal that the hind legs are far more heavily muscled than the forelegs. |
As the brain co-ordinates our nervous activity, so the buttocks co-ordinate our muscular activity. No ape boasts such a muscular monument to compare with ours; and it is a failure more fundamental than his lack of a large brain.14
Certainly, the gluteus maximus is a significant portion of our anatomy. But, did apes "fail" to alter their bodies in this respect? Or did they simply lack the potential for doing so? Perhaps no pure primate had the potential to evolve into a human being by gradual mutation alone. We could, however, have obtained our big rump by other means. One has only to think of a country ham to realize that pigs, too, have powerful buttocks. Perhaps the very first hominid had a large rump as well as many other distinctively human features.
The Spinal Column
Centralization of the spine, another factor facilitating our erect carriage, is not seen in other primates to the extent that it is in humans.15 With the spine shifted toward the center of the body, a larger proportion of the trunk lies to its rear. As a result, the anterior portion is better counterbalanced by the posterior and less effort is required to keep the body erect. In pigs, the spine is more centralized even than our own,16 just as ours is more centralized than an ape's.
| The human sacrum is concave on its anterior face while an ape's is rather flat. The anterior face of a pig's sacrum is markedly concave (405.5,I,35,Fig. 50). |
Centralization of the vertebral column by itself, however, does not account for the ease with which the human body is held erect. Many other modifications of the spine facilitate our bipedality. At the base of the human spine, where the lumbarvertebrae meet the sacrum, is a sharp backward bend known as the lumbo-sacral promontory (see illustration below). The angle formed by this promontory is more acute on the front side of the spine because of subsequent tapering of the sacrum. This configuration causes the sacrum to form the roof of the pelvic cavity in human beings (instead of the rear wall as it does in other primates).17
 |
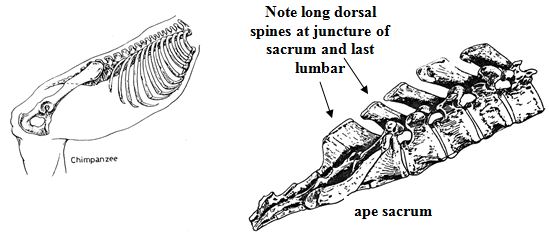 |
 |
More significantly, it brings the base of the flexible portion of the spinal column into a position directly above the hip joints (when viewed from the side). The force applied to the pelvis by the weight of the upper body is directed straight downward through the hip joints and does not tend to rotate the pelvis around those joints. When an ape is fully erect, a vertical line passing through the base of the spine falls behind the hip joints so that a rearward twisting torque is applied to the pelvis. This torque must be countered by constant muscular exertion.
| Barone (55.1,I,439) states that "on thedorsal face of the [pig sacrum] extreme reduction of the dorsal spines is quite characteristic." (translated by E.M. McCarthy) | |
| Krider et al. (280.5,Fig 4-1) provide a photograph of a pig carcass in this position. A lumbo-sacral promontory is clearly visible. |
The dorsal, backward-projecting spine of the uppermost vertebra on an ape sacrum is too long to permit backward flexure of the lowermost lumbar. In human and pig,the spines on the dorsal (back) face of the sacrum are quite short and do not interfere with bending at this point (see illustrations above). But, do pigs have a lumbo-sacral promontory? In anatomical depictions of pig skeletons arranged in the typical quadrupedal pose, no promontory is visible. But if a human being gets down on all fours, then the lumbar region is twisted forward relative to the sacrum, and the promontory disappears.
Perhaps an erect pig would also develop a sharp bend at the base of the spine. Obviously, pigs do not ordinarily stand upright, and I have never seen a drawing showing the configuration of a pig skeleton in such a position. Nevertheless, anyone willing to examine a hanging side of pork will see that a lumbo-sacral promontory is evident. Hanging a halved carcass by the hind leg causes the leg to swing into a position that closely approximates erect human posture. Here, again, porcine anatomy accounts for a human peculiarity.
| Schultz (495.7,77) also points out that "the proportionate length of the lumbar region of man surpasses that of the man-like apes more than [would be] expected from the differences in the lumbar segments." In man the length of the lumbar region is 38 percent of the total length of the trunk, while in the chimpanzee this region is only 22 percent of trunk length. |
The human spine contains more lumbar vertebrae and fewer sacral vertebrae than does the spinal column of any great ape.18 Because sacrals are fused and lumbars are not, the human spine is much more flexible than an ape's. Consequently, we are capable of bending the body backward until it balances over the hip joints (without rotating the pelvis backward). The "small" of the human back is the external evidence of this backward curvature of the lumbars. Pigs have even fewer sacrals19 than do human beings, and they have more lumbars.20 So here, again, humans are intermediate between apes and pigs.
 |
Cervical vertebra (pig). Tracing. |
 |
Cervical vertebra (human) |
 |
Cervical vertebra (ape) |
| Note that the great flexibility of the human neck in comparison with that of apes would make it possible to balance the head, almost regardless of the positioning of the foramen magnum. If the head's ability to swing backward and forward is not limited by long spines on the neck vertebrae, then a balance point will be attainable. | |
| While it is commonly noted that the dorsal spines of the cervical vertebrae slant caudally in Homo, it has also been observed (540.6, 223) that "nonretroverted cervical spinous processes occur frequently in modern Europeans with perfectly normal posture." In the accompanying radiograph tracing (540.6,Fig. 5,223) spines 6 and 7 slope cranially. Pig cervical spines are so short that it is difficult to determine which way they slant except for the long one on the seventh cervical which slants slightly caudally (55.1). |
In all the great apes, the cervical (neck) vertebrae have long dorsal spines — significantly longer than those on their thoracic (ribbed) vertebrae.21 In consequence, ape necks are stiff in comparison with a human being's. Any nodding motion of the head is severely limited.22 Though all cervical spines are long in apes, the fourth and fifth are usually longer than the sixth and seventh. Humans and pigs, on the other hand, have relatively short cervical spines except on the seventh cervical, where the spine is long (but not so long as the thoracic spines).23As a result, humans are better able than apes to adjust the balance of the head by tilting it backward to the equilibrium point.
The seventh human cervical vertebra differs in another respect from those of other primates: it has transverse foramens or "foramina" (see illustration below). These large openings on either side of the spinal canal "are very rarely missing in even the seventh vertebra of Homo sapiens, but in the other primates it is rare to find corresponding foramina in this segment" (Schultz24). In a work on the comparative anatomy of humans and domestic animals, Barone discusses the seventh vertebra, saying it "is not, in general, pierced by a transverse foramen, with the exception of pigs and human beings. In these two cases it always is."25
 |
Seventh cervical vertebra (human) |
Pelvis and Coccyx
| Schultz (495.06,429) states that "In the macaque and gorilla, as well as in the other monkeys and apes examined for these conditions, there is no fixed, bony structure opposite the pubic bones, as exists in man in the form of the lower part of the sacrum. In the former, therefore, the sacrum interferes not nearly so much with the passage of the fetus to be born, as in the latter." The obstruction of the birth canal by the sacrum in human beings reflects the shortness of the human pelvis in comparison with the simian. This shortening can be accounted for by the fact that pigs have a very short pelvis. A small pelvic opening does not interfere with parturition in swine because their newborns are relatively small in comparison with those of primates. |
At the opposite end of the spine are the coccygeal vertebrae which together form the coccyx, or tail bone. Adolph Schultz observes that these vertebrae are fused in chimpanzees, a lack of flexibility he terms "puzzling."26
The human pelvis and birth canal are smaller than those of apes.28 Moreover, the sacrum and coccyx curve inward in humans to make a sharp-pointed obstacle that must be negotiated by an emerging infant.29 In apes there is no curvature (see illustration above), which leaves the birth canal unobstructed.30With their constricted birth canals, human females experience far more difficulty in delivery than do their simian counterparts. "Parturition in the great apes is normally a rapid process," according to primatologist A. F. Dixson, who further states that
Gorillas, orangutans and chimpanzees typically give birth in less than one hour and in most cases there is little sign that parturition is imminent … The rapidity with which the great apes give birth correlates with the fact that the head of the newborn is remarkably small in comparison to the female's pelvic canal. In human females, by contrast, labor may be prolonged and the baby's large head is often turned sideways to facilitate its passage through the canal.31
| "The antero-posterior diameter extends from the tip of the coccyx to the lower part of the pubic symphysis," says Gray (220.1,267). "It varies with the length of the coccyx, and is capable of increase or diminution, on account of the mobility of that bone." |
Turning of the head occurs in Homo sapiens because the pelvis is so short that the birth canal is wider than it is high (unlike other primates).32 In humans, the height (antero-posterior diameter) of the birth canal depends on the length of the coccyx and, specifically, on how closely the tip of the coccyx approaches the front wall of the passage.
| Contrary to popular belief, it is not merely the human head, but the entire body that is larger than that of any ape at birth (460.5,73,Table 3). Even the gorilla does not catch up with human babies in size until the second year (460.5,74,Fig. 10). This may be a manifestation of heterosis. In proportion to body size, the head of a new-born ape is as large as that of a human being. In both cases, the brain composes about 12 percent of body weight (188.7). |
The human coccyx, then, ought to be relatively short, since the human neonate is larger than any newborn ape.
And yet, "man is distinct among higher primates by possessing the largest average number of coccygeal vertebrae, i.e., by having been so far affected least by the evolutionary trend to reduce the tail" (Schultz33). "In the human coccyx there may be as many as six elements, in the anthropoids there are quite commonly only two. The anthropoids have gone further than man in the reduction of the tail" (Jones34). This longer "tail" is difficult to account for in terms of natural selection. With respect to reproduction, it is clearly a negative factor. Nor does it have any evident utility in other respects.
| << A similar conclusion was reached by Straus: "The human ilium would seem most easily derived from some primitive member of a pre-anthropoid group, a form which was lacking in many of the specializations, such as reduction of the iliac tuberosity and anteacetabular spine and modification of the articular surface, exhibited by the modern great apes. I wish to emphasize here that the anthropoid-ape type of ilium is in no sense intermediate between the human and lower mammalian forms. Its peculiar specializations are quite as definite as those exhibited by man, so that it appears very unlikely that a true anthropoid-ape form of ilium could have been ancestral to the human type." Quoted in (495.06,431). |
distinguishing characters of the human ilium [upper portion of pelvis] are so numerous and in most instances so very pronounced, whereas the ilia of all the anthropoid apes show so many basic similarities, that no theory which derives man from a gorilla-chimpanzee stock can readily account for these conditions.35
The most obvious difference is the shortness of the human ilium. The pelvis of an ape is about half again as long as a human's (as a percentage of body length) and closely approaches the last rib36 (in the great apes, Schultz (495.7,76) notes that the iliac crest approaches the last ribs "far more closely than in any other primates"). A pig has a short pelvis and a wide gap between pelvis and rib cage, just as we do. The upper blades of the pelvis run from side to side in apes but turn towards the front in humans.37 They also turn forward in pigs.38
Lower Extremities
All nonhuman catarrhine primates have longer arms than legs.39 The reverse is the case in humans. But pigs, like humans, have longer hind limbs than forelimbs.40 The femur(thighbone) is the largest bone of the body. Paleoanthropologists distinguish the femur of a hominid from an ape's in several ways. On the front of the lower end of the femur in humans and apes, the patellar groove forms a track for the kneecap. In apes, this groove is relatively shallow and its medial lip is more prominent than the lateral.41 But in humans42 (and in pigs43) this groove is deep and the lateral lip is the more elevated of the two.
| Physical anthropologists often note that the intercondylar fossa (or notch) is deeper in Homo sapiens than in pongids (325.5,308; 445.5,282; 468.2). Barone (55.1,I,693) describes the porcine intercondylar notch as "très profunde" (very deep). |
Also, on the distal (lower) end of the femur are twocondyles. In Homo, these condyles are of approximately equal size. In pongids the medial one is markedly larger than the lateral,44 but in pigs the femoral condyles are almost exactly equal in size.45Human femoral condyles also differ in shape from those of other primates.
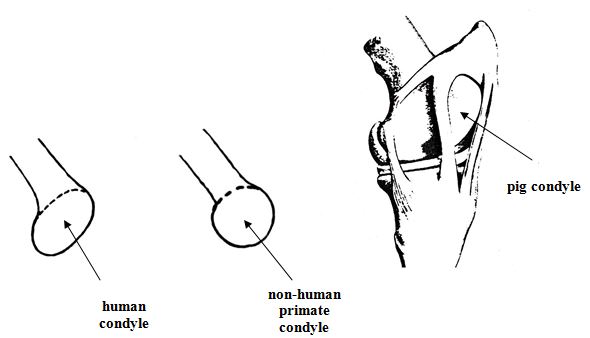 |
Lateral views of femoral condyles in humans, non-human primates and pigs |
| Lovejoy (325.5,310) suggests that a prominent lateral lip is an adaptation "directly related to a valgus knee position produced by a high bicondylar angle." The horse, however, has a very high bicondylar angle (that is, it is quite knock-kneed) and yet the medial lip is much more prominent than the lateral. Knock-knees, then, do not always result in a prominent lateral lip. | |
| Approximate measurements I have taken from anatomical drawings (405.5,I,89) give a bicondylar angle of about 15 degrees for the pig femur, which suggests that pigs are more knock-kneed than most human beings. |
Quadrupedal primates are bowlegged, especially the anthropoid apes.48 Human beings, however, are typically knock-kneed.49 Preuschoft50 follows Kummer51in suggesting that our knock-kneed stance is an adaptation facilitating bipedal posture, and bowlegs, to quadrupedal posture. But the domestic quadrupeds (dog, horse, cow, pig, etc.) are consistently knock-kneed.>
In pigs (and most other domestic animals), the femoral condyles rest on crescentic menisci that are connected to the tibia (shinbone) in the same way as in humans.52 This configuration is significant because, as Tardieu53points out, Homo sapiens is the only primate having a "crescent-shaped [lateral] meniscus with two tibial insertions." In fact, in the vast majority of catarrhine primates (including the chimpanzee and gorilla) the lateral meniscus is ring-shaped. In the tibia itself the most prominent difference is the presence of a long malleolus medialis in nonhuman primates.54
| Romer (470.4,273) remarks that, in artiodactyls, "the astragalus is the most characteristic bone in the skeleton, for it has not only a rolling pulley above, but an equally developed lower pulley surface [articulating with thecalcaneus]. This type of articulation gives very great freedom of motion to the ankle for flexion and extension of the limb and a potential springing motion, but limits the movement to a straight fore-and-aft drive even more strictly than is the case in perissodactyls [odd-toed ungulates]." |
We find another human distinction in the foot, in the joint between the heel bone (calcaneus) and the anklebone (astragalus). Szalay and Delson note that one feature distinguishing hominids from apes is the "loss of [the] ancestral helical astragalo-calcaneal articulation, reducing the possible range of movements in this joint."56 In apes the articulation is "helical" because the joint allows rotation of the foot in a plane parallel to the ground. In Homo sapiens, this joint is more like a hinge. It allows only flexion and extension.57 A pig, too, has a hinge-type articulation between the calcaneus and the astragalus.
| The upper surface of the talus in human beings is level from side-to-side so that it is parallel to the base of the foot (542.8,52). In chimpanzees (ibid.) this surface lies at an angle so that a perpendicular to it passes through the lateral side of the sole of the foot — this angulation affects the way apes walk when upright. In taking a step, all of the pressure is placed on the outside edge of the foot. Instead of rising on its toes at the end of a step, an ape rolls the pressure point forward along the side of the foot in a rocking motion. According to Sisson (525.3,184,Fig. 197) the porcine tibiotarsal joint is level, as it is in human beings. |
The proportions of the human foot are also peculiar for a primate. Duckworth notes that the human heel bone is longer than that of apes.58 Baba found that the length of the third metatarsal bone exceeded the length of the calcaneus in all primates in his survey — except in humans, in which the calcaneus is slightly longer(the third metatarsal connects the middle toe with the ankle and composes most of the length of the foot between the ankle and ball of the foot).59Our high ratio of calcaneus to metatarsal makes it easier for us to bear the body's weight on the ball of the foot (as we do each time we take a normal step), because the forepart of the foot and the heel bone can be thought of as two ends of a lever having the ankle as a fulcrum. As in humans, the heel bone is a bit longer than the third metatarsal in domestic pigs.60
| The fact that our toes are shorter than our fingers can be accounted for under the hybrid hypothesis by the fact that in chimpanzees the toes are markedly shorter than the fingers. |
Our fingers and toes, are short compared to those of apes.61 Our metacarpal bones and phalanges are shorter than a chimpanzee's (not just in relation to the overall length of the hand, but absolutely).62 This shortening can be explained by referring to the anatomy of pigs: their digits are even shorter and stubbier, than our own (which, of course, is the case for most quadrupeds).
| Shrewsbury and Sonek (510.6,237) feel that the difference between human nail phalanges and those of other primates is so marked that a distinction in terminology is called for, saying, "For humans we reserve the diagnostic term ungual tuft; for non-human primates the term ungual tuberosity is to be employed … [because] the roughened development of the volar aspect of a broadened ungular tuft, characteristic of humans, is not evident in the prevailingly conical ungual tuberosities of the other primates." While it does seem that a distinction in terminology is called for, it makes more sense to use a new term in connection with nonhuman primates instead of with human beings, because the term ungual tuberosity was originally used in describing humans. Moreover, no tubercle being present in these animals, the choice of the termtuberosity seems inappropriate. |
Lastly, consider the ungual tuberosities. These small, hoof-shaped processes tip the bones (nail phalanges) that underlie the nails of our fingers and toes (see illustration below). Nonhuman primates do not have such processes. "When comparing the nail phalanges of apes to those of man, a pronounced slenderness of the former can be observed. If the impressive strength of pongid hands is taken into consideration, this is surprising" (Preuschoft63).
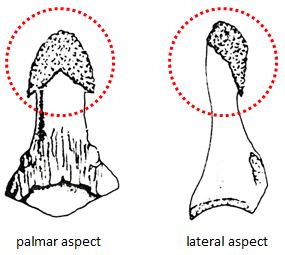 |
Human distal phalanx (ungual tuberosities circled in red). |
Convergence or Relationship?
| Elsewhere on this website, some of the problems with thinking in terms of homology and analogy are considered at length. Access this discussion >> |
Our hypotheses have accounted for a number of traits in Homo. From the standard neo-Darwinian perspective, it is hard to understand why the parallels between human being and pig should be so extensive. Biologists call the existence of similar traits in animals that they consider to be distantly related analogy. They say analogy is found when animals live under similar conditions or have similar habits. The same needs in each case are supposed to cause structures of similar function to develop during the course of evolution. But when the organisms under consideration are considered to be closely related, such features are termed homologous. Homologous features are usually judged to be so when the similarities are numerous and extend to detail. As Dobzhansky et al. put it, "Examination of the structure of convergent features usually makes it possible to detect analogy because resemblance rarely extends into the fine details of complex traits."65
In this section we have considered one complex trait (bipedality) in a fair amount of detail. Any attempt to account for these details in terms of natural selection seems inadequate. It is difficult to see what “selective pressures” could have caused human beings and pigs to “converge” in so many different respects. Under neo-Darwinian theory, to explain most of the human features that we have just discussed, we have to posit pressures selecting for bipedality (some human features — long tail bone and ungual tuberosities — cannot be explained in this way). But pigs are quadrupeds. How will we account for the fact that they, too, have these features? Perhaps it is all just a coincidence, but after a certain point coincidence begins to assume the color of relationship.
| A list of traits distinguishing humans from other primates (and shared with pigs) DERMAL FEATURES Naked skin (sparse pelage) Panniculus adiposus (layer of subcutaneous fat) Panniculus carnosus only in face and neck In "hairy skin" region: - Thick epidermis - Crisscrossing congenital lines on epidermis - Patterned epidermal-dermal junction Large content of elastic fiber in skin Thermoregulatory sweating Richly vascularized dermis Normal host for the human flea (Pulex irritans) Dermal melanocytes absent Melanocytes present in matrix of hair follicle Epidermal lipids contain triglycerides and free fatty acids FACIAL FEATURES Lightly pigmented eyes common Protruding, cartilaginous mucous nose Narrow eye opening Short, thick upper lip Philtrum/cleft lip Glabrous mucous membrane bordering lips Eyebrows Heavy eyelashes Earlobes FEATURES RELATING TO BIPEDALITY Short, dorsal spines on first six cervicalvertebrae Seventh cervical vertebrae: - long dorsal spine - transverse foramens Fewer floating and more non-floating ribs More lumbar vertebrae Fewer sacral vertebrae More coccygeal vertebrae (long "tail bone") Centralized spine Short pelvis relative to body length Sides of pelvis turn forward Sharp lumbo-sacral promontory Massive gluteal muscles Curved sacrum with short dorsal spines Hind limbs longer than forelimbs Femur: - Condyles equal in size - Knock-kneed - Elliptical condyles - Deep intercondylar notch at lower end of femur - Deep patellar groove with high lateral lip - Crescent-shaped lateral meniscus with twotibial insertions Short malleolus medialis Talus suited strictly for extension and flexion of the foot Long calcaneus relative to foot (metatarsal) length Short digits (relative to chimpanzee) Terminal phalanges blunt (ungual tuberosities) Narrow pelvic outlet ORGANS Diverticulum at cardiac end of stomach Valves of Kerkring present in small intestines Mesenteric arterial arcades Multipyramidal kidneys Heart auricles level Tricuspid valve of heart Laryngeal sacs absent Vocal ligaments Prostate encircles urethra Bulbo-urethral glands present Os penis (baculum) absent. Hymen Absence of periodic sexual swellings in female Ischial callosities absent Nipples low on chest Bicornuate uterus (occasionally present in humans) Labia majora CRANIAL FEATURES Brain lobes: frontal and temporal prominent Thermoregulatory venous plexuses Well-developed system of emissary veins Enlarged nasal bones Divergent eyes (interior of orbit visible from side) Styloid process Large occipital condyles Primitive premolar Large, blunt-cusped (bunodont) molars Thick tooth enamel Helical chewing BEHAVIORAL/PHYSIOLOGICAL Nocturnal activity Particular about place of defecation Good swimmer, no fear of water Extended male copulation time Female orgasm Short menstrual cycle Snuggling Tears Alcoholism Terrestrialism (Non-arboreal) Able to exploit a wide range of environments and foods RARE OR ABSENT IN NONHUMAN PRIMATES: Heart attack Atherosclerosis Cancer (melanoma) |
| Why genetic data cannot be used to evaluate the hypothesis In connection with the hypothesis that human origins can be traced to a hybrid cross, it's important to realize that in most mammalian hybrid crosses, the male hybrids are usually more sterile than are the females. This fact means that breeders working with hybrids typically mate fertile females with one of the two parents (that is they "backcross" them). They do not, as a rule, produce new breeds by breeding the first-cross hybrids among themselves. Often, even after a backcross, the resulting hybrids are still fertile in only one sex. So repeated backcrossing typically occurs. However, after a sufficient number of backcrosses, fertile hybrids of both sexes are often obtained and the new breed can thenceforth be maintained via matings among the hybrids themselves. Repeated backcrossing tends to be more necessary in cases where the parents participating in the original cross are more distantly related and genetically incompatible. So one expects also, in the case of new types of organisms arising via natural hybridization, for backcrossing to be the usual route to fertility and reproductive stability. And the same would hold in the specific case of humans arising via hybridization. However, consider the effect of such repeated backcrossing on the human genome. The reader may not be familiar with the phenomenon of gene conversion, but its effect on hybrids during backcrossing is to quickly homogenize gene sequences. To understand why this is the case, consider the effects of backcrossing on hybrid DNA.
In the figure above, note that in either of the parental (incoming) double strands each nucleotide in one strand is properly paired with its complementary nucleotide in the other strand, A is always paired with T, and C is always paired with G. So the paired double DNA strand from one parent might look like this: AGTTCCGACGCG TCAAGGCTGCGC while the strand from the other parent might look like this: AGCTCCGACGCG TCGAGGCTGCGC In each of these two double strands all nucleotides are paired with the complementary nucleotide (A always with T, and C always with G). But when one double strand is compared with the other it's clear that they differ at the third nucleotide base position. In the first double strand the nucleotide base pair is T-A, while in the second it is C-G. Thus, when these strands associate with their new partner strands after passing through the Holliday junction in meiosis, the two resulting double strands will be: AGTTCCGACGCG TCGAGGCTGCGC and AGCTCCGACGCG TCAAGGCTGCGC So T is paired with G in the first outgoing strand and C is paired with A in the second outgoing strand. Gene conversion converts each such mismatched pair into a matched one by replacing one of the two nucleotides with the complement of the other nucleotide (which remains unchanged). Experimental results suggest that the mechanism chooses at random which of the two nucleotides to replace so that the nucleotides derived from either parent survive at equal rates. So, in an given case of backcrossing, suppose the genomes of the original parents A and B, which produced the first-cross hybrids, differed at one nucleotide position in five (20%). Then the DNA in gametes of the hybrids would differ from A at one position in ten and from B at one position in ten. That is, the first cross hybrid's gametes would be right in the middle between A and B with respect to gene sequence. However, after one backcross to A, and the resulting gene conversion during meiosis in the backcross hybrids, the gametes produced by backcross individuals would differ from A at only one position in twenty (5%). And gametes produced by second backcross hybrids would differ at only one position in forty (97.5% similarity). It's clear then that it rapidly becomes quite difficult to distinguish, on the basis of nucleotide sequence data, the backcross hybrids from the pure parent A to which backcrossing has occurred. Chimpanzees and humans are about 98% similar in terms of their nucleotide sequences. |
Subscribe to:
Posts (Atom)
Popular Posts
-
This was one big mother fucker. The bomb was built and eventually tested on October 30, 1961, on the west coast of Novaya Zemlya Island (A...
-
Radial Engines Radial engines are used in aircrafts having propeller connected to the shaft delivering power in order to produce thrust its ...
-
Source Every once in a while archaeologists (and sometimes regular Joes) make some remarkable discoveries. Stunned, they are often unable to...
-
" Comet Elenin, Earth, and the Sun were in exact alignment last year on February 27th. That is when the Chile Earthquake happened, and ...
-
 Thanks to the internet, there are numerous pictures and videos on freaky sea creatures out there. The main point of this post is not to debu...
Thanks to the internet, there are numerous pictures and videos on freaky sea creatures out there. The main point of this post is not to debu... -
Source Bigfoot, the Loch Ness monster, and El Chupacabra are all creatures that are regulated to myth. They have been ignored by mainstream ...
-
Source by Steve Quayle Ancient India Puranas and their history are NOT Mythology. it comes from ancient Vedic civilization .What caused ...
-
From Cracked.com By Jake Slocum Oct 02, 2008 2,573,979 views 0 digg The world is a magical place, full of mysteries science may never unde...
-
Source by jewditzsue A cryptid is a creature who may be thought by some people to exist, but is not recognized by the scientific community. ...








